Предисловие к русскому изданию
Предполагая, что очерк естественной истории воспаления может найти читателей в России, я решил издать русский перевод моих лекций, читанных в прошлом году в Пастеровском институте.
Перевод, сделанный П. Циклинской и О. Мечниковой, был просмотрен мною, так что ответственность за него падает на меня. Передавая его на суд читателя, я должен прежде всего предупредить, что предлагаемый мною труд не имеет претензии быть полным трактатом о воспалении. Некоторые стороны этого вопроса, как, например, этиология нагноения, были преднамеренно исключены из этого очерка, чтобы не слишком усложнять его.
Главная цель моего труда состоит в том, чтобы установить прочную связь между патологией и зоологией, или, правильнее, биологией вообще.
Подобно тому как в прежние времена сравнительная анатомия ограничивалась лишь изучением человека и высших животных, в медицине и до сих пор еще упорно игнорируются все патологические явления низших животных. А между тем эти последние, представляя условия наиболее простые и первобытные, дают, так сказать, ключ к пониманию сложных патологических явлений, входящих в область медицины.
Изучение воспалительного процесса с сравнительно-патологической точки зрения дает нам возможность проникнуть гораздо глубже в сущность этого явления.
* Опубликованы в 1892 г. по-французски отдельной книгой в Париже и в том же году в русском переводе. — Ред.
263
В биологической теории воспаления, которую я провожу в этом очерке, очень часто упоминается о фагоцитах. Я должен, однакоже, предупредить читателя, что я в этом сочинении не даю полного изложения теории фагоцитов, так как некоторые стороны ее, как, например, вопросы об иммунитете, о выздоровлении и атрофии, должны быть изложены особо.
За весьма малыми исключениями, лекции эти изданы в том виде, как они были прочитаны мною. Изменений против французского подлинника сделано не было.
Ил. Мечников Париж, 19/31 марта 1892.
ЛЕКЦИЯ ПЕРВАЯ
Инфекция есть борьба между двумя организмами. — Пример Sphaerophrya. — Сравнительная патология как ветвь зоологии. — Основные элементы этой науки. — Воспаление. — Обзор главных теорий этого явления. — Современное положение вопроса. — Необходимость сравнительного метода для изучения воспаления
Главнейшие патологические процессы исследуются обыкновенно на позвоночных, да и то начиная с лягушек, т. е. с животных уже очень сложных. Этим объясняется крайняя трудность изучения и анализа патологических явлений. Но задача может быть значительно облегчена, если мы обратимся к низшим животным, у которых все процессы гораздо проще. Исходя из такой точки зрения, я и решаюсь в качестве зоолога прочесть эти несколько лекций.
Нет надобности приводить специальные доказательства в подтверждение того, что болезнь и патологические процессы следуют тем же законам эволюции, как сам человек и высшие животные. Все организмы, начиная с самых низших, подвержены инфекционным болезням, вызываемым различными паразитами. Совершенно естественно предположить, что паразитизм производит целый ряд определенных болезненных явлений в зараженном организме и вызывает в то же время реакцию с его стороны.
Исследуя животных или растения, мы находим, что очень многое в их организации приспособлено к нападению или защите. Твердые покровы рака, скорлупа моллюсков, зубы позвоночных и многие другие органы — все это охранительные средства животных в их постоянной борьбе. Одно перечисление подобных органов потребовало бы полного изложения сравнительной анатомии.
Не трудно доказать, что активное нападение легко превращается в инфекцию. Спустимся в мир низших существ и остановимся на биологических отношениях некоторых видов инфузорий.
Между этими животными есть группа сосущих — ацинеты. Они снабжены присосками, направленными в разные стороны и служащими для нападения на инфузорий других видов и для высасывания всего их содержимого (рис. 1.). Большинство этих ацинет прикрепляется к различным подводным предметам
265
и, играет роль настоящих хищников. Наблюдая их образ жизни, можно легко объяснить многие стороны их организации, приспособленной к активной борьбе с другими инфузориями; сопротивление последних также совершенно ясно.
Между ацинетами есть очень мелкие представители. Вместо того чтобы быть прикрепленными к подводному предмету и привлекать к себе добычу, они, напротив, свободны и, выбирая более крупных инфузорий, сами прикрепляются к ним. Эти маленькие ацинеты проникают сквозь покровы своей добычи, внедряются внутрь ее и ведут в ней паразитический образ жизни (рис. 2).
Пример этот показывает нам, что очень близкие организмы — два вида одного и того же рода (Sphaerophrya magna и Sph.
paramaeciorum) — могут, с одной стороны, быть нападающими хищниками, а с другой — паразитами, производящими настоящую инфекцию. Если явления нападения и защиты составляют предметы зоологического исследования, то и столь близкие им явления инфекции и сопротивления ей входят в ту же область исследования. Между этими явлениями только та разница, что активная борьба между животными бросается в глаза наблюдателю, и потому естествоиспытатели давно обратили на нее внимание, тогда как более скрытые явления инфекции были наблюдаемы только очень редко и недостаточно 155.
Итак, одна из отраслей зоологии должна быть посвящена изучению многочисленных приспособлений животного организма для проникновения в другое животное и для жизни в нем. С другой стороны, и главным образом, она должна изучать органы защиты и явления реакции в организме, подверженном нападению со стороны паразитов.
Таким образом должна возникнуть отрасль общей зоологии, т. е. сравнительная патология животных, которая будет во многих отношениях отличаться от ныне существующей сравнительной патологии. В то время как эта последняя, основанная главным образом ветеринарами, применяется исключительно к высшим животным, а именно к позвоночным, настоящая сравнительная патология должна обнимать весь животный мир в его целом и изучать его с самой общей биологической точки зрения.
Основные элементы сравнительной патологии, изучаемой с этой точки зрения, были заложены уже приблизительно треть
266
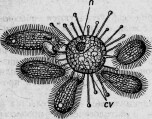
Рис. 1. Sphaerophrya magna (no Мопа)
века тому назад: почти одновременно, в 1857 и 1858 гг., Дарвином и Уолласом была обоснована научным образом теория естественного подбора; затем Пастером — биологическая теория брожения и Вирховым — целлюлярная патология. Первая из этих теорий, служащая в настоящее время основанием для всякого биологического исследования, доказала преемственность развития организованных существ и объяснила явление целесообразности. Она констатировала, что одни полезные признаки упрочиваются борьбой за существование, вредные же устраняются естественным подбором.
Биологическая теория брожения, созданная Пастером, благодаря его открытию фермента молочнокислого брожения в 1857г. и фермента масляного брожения в 1861 г., — сразу указала путь для объяснения причин инфекций. Давен получил вследствие этого возможность выдвинуть забытое открытие микроба сибирской язвы и тем дать точку отправления патологической бактериологии.
Наконец, выяснение Вирховым важной роли клеток организма в патологических процессах послужило третьим неизбежным звеном в цепи этих биологических теорий, необходимых для построения настоящей сравнительной патологии 156.
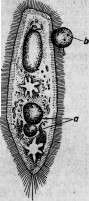
Рис. 2. Sphaerophrya paramaeciorum: a — две паразитические Sphaerophrya; b — такой же индивидуум, прикрепленный к поверхности парамециума.
Но несмотря на то, что основы этой науки были уже давно установлены, мы и в настоящее время еще недостаточно подготовлены для изучения вопросов общей патологии с сравнительной точки зрения. Доказательством этого могут служить господствующие учения патологии, относящиеся к наиболее важным болезненным явлениям.
Возьмем, например, воспаление, которое есть, как всем известно, явление, преобладающее во всей патологии. Познакомимся сначала с результатами, добытыми относительно этого явления обычными методами, и затем посмотрим, не удобнее ли перенести . вопрос о воспалении на почву сравнительной патологии. Не имея возможности сделать подробного изложения всех теорий воспаления, ограничимся теми, которые имели наибольшее влияние в патологии и преподаются большей частью в настоящее время.
Очень долго внимание врачей было обращено главным образом на самый внешний признак воспаления — красноту. Вследствие этого на прилив крови смотрели как на существеннейшее явление воспаления и даже отождествляли их. Теории воспаления сводились поэтому к анализированию гиперемии и ее
267
причину искали или в сосудодвигательном параличе (паралитическая теория), или в спазмодическом сокращении поврежденных артерий, связанном с приливом крови к окружающим областям (спазмодическая теория). Но скоро должны были заметить, что одна только гиперемия неспособна обусловить настоящее воспаление. Часто наблюдается временная гиперемия, иногда даже довольно продолжительная, но без всякого экссудата, неизбежного при типичном воспалении. Поэтому для объяснения «опухоли» придумали теорию, по которой поврежденная область имеет притягательную силу для крови. Эта теория получила более определенный смысл благодаря учению Вирхова, а именно, что клетки воспаленной области обладают усиленной способностью питания и размножения, обусловливающими развитие большого количества клеток экссудата на счет элементов самой поврежденной ткани. По этой теории гиперемия представляется явлением подчиненным и вполне второстепенным.
Установление Конгеймом факта, что клетки воспалительного экссудата происходят из белых кровяных телец, позволило впервые решить точным образом один из главных вопросов воспаления. Установив незыблемо этот факт, Конгейм присоединился к теории Самуэля, по которой самая суть воспаления заключается в болезни сосудов, вызванной известным раздражителем. Воспаленные сосуды, сделавшись менее прочными, дают возможность пассивному выхождению жидкостей и кровяных шариков. Эти экссудативные части скопляются в месте наименьшего сопротивления, вследствие чего и образуется воспалительная опухоль *.
По теории Самуэля и Конгейма, ткани поврежденной области, так же как и гиперемия и явления со стороны сосудодвигательных нервов, имеют только второстепенное значение при воспалении. Хотя некоторые пункты этой теории встретили более или менее серьезные возражения, но она принята большинством современных патологов. Это учение часто принимается ими только в общих чертах, причем последователи его настаивают в то же время на возможности явлений, происходящих в самой поврежденной ткани и в сосудодвигательных нервах. Не имея возможности подвести все эти явления к общему началу, они довольствуются простым перечнем изменений, вызванных воспалением в тканях и в сосудодвигательной системе.
Циглер **, автор наиболее распространенного в настоящее время руководства патологической анатомии, признается в главе «Об определении воспаления» в невозможности дать этому
* Опухоль — слово, применяемое здесь Мечниковым в устарелом его понимании. В настоящее время строго различают опухоль — разрастание ткани организма и простую припухлость, отек, вызываемый скоплением жидких элементов в воспаленном участке. — Ред.
** Lehrbuch der pathol. Anat, т. I, изд. 6, 1889, стр 186.
268
явлению точной формулировки. Вот что он говорит по этому поводу: «Понятие «воспаление» включает в себя целый ряд явлений, происходящих отчасти в сосудистой системе, отчасти в тканях; эти процессы связаны между собой различным образом. Так как мы, следовательно, имеем дело не с одним отдельным явлением, то краткое и точное определение «воспаления» становится невозможным. Если даже взять только характерные для воспаления явления, например, те, которые происходят в сосудистой системе, то и тогда определение их не исчерпало бы понятия о воспалении». После этого Циглер ограничивается простым описанием изменений, вызванных воспалением.
По мнению Реклинггаузена *, «невозможно в настоящее время определить «primum movens»** воспаления, исходную точку изменений, т. е. место первичного повреждения». Таким образом, и ему остается одно подробное и тщательное описание явлений воспаления.
Определение воспаления, сделанное Корнилем и Ранвье ***, сводится к простому перечню явлений этого патологического процесса. Они определяют его как «ряд явлений, наблюдаемых в тканях или органах. Аналогичные им явления можно произвести искусственно действием физического или химического раздражителя».
Для упрощения столь сложного и трудного вопроса давно уже стали искать такую часть организма, которая была бы лишена сосудов и где можно было бы наблюдать изолированно воспалительные явления. Внимание было поэтому сосредоточено на хрящах, сальнике и, главным образом, на роговой оболочке. Из изменений, наблюдаемых в клетках этих тканей, был сделан вывод, что участие сосудов при воспалении не является необходимым. В бессосудистых тканях воспалительные явления выражались в размножении местных клеточных элементов и в возвращении их к «эмбриональному состоянию».
В своих исследованиях роговой оболочки Конгейм показал, что искусственный кератит связан с вмешательством со стороны сосудов, причем белые кровяные шарики мигрируют в воспалительный очаг то с краев роговой оболочки, то из конъюнктивального мешка. Эти результаты показали в то же время несостоятельность попыток выделить влияние сосудов при воспалении даже в таких органах высших животных, где они всего более отсутствуют.
Затем стали стремиться ввести в цикл воспалительных явлений изменения, происходящие внутри самих клеток, и восстановили теорию паренхиматозного воспаления Вирхова. Значительно расширив, таким образом, область воспаления, старались
* Handbuch d. allgemeinen Pathologie des Kreislaufes, 1883, стр. 198. ** Первый толчок (лат.). — Ред. *** Manuel d'histologie pathologique, т. I, изд. 2, 1884, стр. 94.
269
включить в нее и явления острой дегенерации клеток, как это было сделано Бро* в последние годы.
Итак, видно, что главная забота патологов сосредоточивалась в последнее время на определении взаимной роли сосудистой системы и местных клеточных элементов тканей.
Спор о происхождении воспалительных клеток из вышедших белых кровяных шариков или из местных клеточных элементов был возобновлен благодаря открытию кариокинетических явлений.
Это дало возможность решить многие вопросы развития и происхождения клеток.
В последнее время возникла полемика между Гравицом **, учеником Вирхова, и Вейгертом ***, последователем Конгейма.
Первый доказывает, что большая часть гноя образуется за счет соединительнотканных клеток; второй поддерживает основную тезу своего учителя о происхождении воспалительных клеток из лейкоцитов, эмигрирующих сквозь стенки сосудов.
Открытие кариокинезиса дало возможность точно констатировать очень частое деление местных клеток воспалительного очага. Последователи Вирхова сделали из этого вывод о роли местных элементов в образовании воспалительной опухоли; защитники же воззрений Конгейма видели в делении клеток исключительно акт восстановления данного органа от первичного повреждения. Склоняясь все более и более к этому последнему воззрению, стали различать в воспалении две категории явлений: во-первых, воспаление в непосредственном значении слова, т. е. повреждение стенок сосудов и другие болезненные явления, вызванные раздражителем; во-вторых, восстановление, заключающееся в регенерации недостающих тканей и в образовании рубца. Розер **** пошел дальше всех в этом направлении. Он доказывает, что воспаление есть болезнь, вызванная микробами, а выздоровление слагается из явлений восстановления.
Розер считает даже «немыслимым дать общее определение воспалению, пока под этим именем соединяют самые разнородные явления, как инфекционные болезни и процессы выздоровления».
Но рядом с этим взглядом на воспаление давно уже возник другой в противоположном направлении. Вместо того, чтобы строго разделять две категории воспалительных явлений, старались объяснить их в целом как полезную реакцию против данного вредного деятеля. По этому учению, не только регенерация и зарубцевание, но и явления первичного воспаления, эмиграция лейкоцитов и изменение стенок сосудов служат восстановлением от повреждения.
* Etude sur I'nflammation, Paris, 1888, стр. 34. ** Deutshe med. Wochenschr., 1889, № 23. *** Fortschritte der Medicin, 1889 № 15 и 16. **** Entzundung und Heilung, Leipzig, 1886, стр. 9, 11 и сл.
270
К этой теории, высказанной Заксом в очень определенной форме более 50 лет назад, возвращались в разные периоды времени. Поддерживаемая в общих чертах Бухнером *, она была развита в последнее время Нейманом **. Он утверждает, что настоящее воспаление происходит исключительно в случаях первичного повреждения тканей, и старается установить следующее определение воспаления: «под этим именем мы должны рассматривать ряд местных явлений, развивающихся вследствие первичных повреждений тканей (Laesio continui или некроз) и приводящих к их восстановлению» (1. с., стр. 363).
Из этого очерка современного учения о воспалении видно, что это явление, будучи в высшей степени сложным и изменчивым в своих проявлениях, не может быть достаточно изучено общепринятыми методами, несмотря на все старания исследователей. Поэтому понятно, почему некоторые ученые, как, например, Тома ***, предлагали полное устранение термина «воспаление».
Несмотря на частые попытки упростить исследование, исключая некоторые факторы из сложного процесса воспаления, до сих пор удалось получить положительные результаты только относительно воспалительного жара.
Изучая воспаление у лягушек, легко можно было убедиться, что здесь идет дело о настоящем воспалении, несмотря на отсутствие одного из четырех основных факторов его — dolor, calor, rubor, tumor 157 — повышения температуры. Воспалительный характер наблюдаемых явлений здесь настолько резок, что никому не пришло в голову возражать против термина «воспаление». Между тем в данном случае отсутствует именно повышение температуры, так что слово «воспаление» не применимо в этимологическом смысле.
К лягушке обратились только вследствие громадного преимущества этого животного с чисто технической точки зрения, не подозревая вовсе, что в то же время этим самым прибегли к сравнительно-патологическому методу. Для того чтобы выделить все несущественные факторы воспаления, надо следовать по тому же пути и сойти еще ниже по лестнице животных существ.
Мы уже видели, что невозможно вызвать воспаление без вмешательства сосудов, потому что у высших животных, даже в наиболее изолированных тканях, нельзя вполне устранить роли сосудистой системы. Поэтому для получения положительных результатов надо обратиться к многочисленной группе беспозвоночных; между ними есть много представителей, вполне лишенных сосудов.
* Prophylactische Therapie der Lungentuberkulose, 1882. ** Ueber den Entzundungsbegriff. Beitrage zur pathol. Anat. (Ziegler), т V, 1889, стр. 347.
*** Berliner klin. Wochenschr., 1886.
271
Сравнительный метод оказал уже свои услуги не только в области естественных наук в строгом смысле слова, но даже в изучении наиболее сложных вопросов. Так, психология почерпнула много полезных данных из изучения психической деятельности наиболее простых животных... Одна только патология до сих пор игнорирует сравнительный метод, несмотря на то, что она имеет дело с очень сложными во многих отношениях явлениями. Именно она должна бы всего более черпать в этом методе для расширения поля своих исследований.
Можно следующим образом формулировать вопрос, поставленный на эту почву: обусловливается ли воспаление у низших позвоночных (амфиоксус) и у беспозвоночных теми же факторами, которые вызывают его у человека и высших позвоночных? Необходимо ли для воспаления присутствие сосудистой системы, или же оно может совершаться также у животных, лишенных сосудов? Какая в последнем случае роль нервной системы? Необходимо ли животному для воспаления обладать известным количеством дифференцированных органов, или для этого достаточно, чтобы оно состояло из скопления недифференцированных клеток? Встречается ли в растительном мире что-нибудь аналогичное воспалению? Обнаруживают ли одноклеточные организмы воспалительные явления?
В следующих лекциях мы рассмотрим отдельно каждый из этих вопросов.
ЛЕКЦИЯ ВТОРАЯ
Подвержены ли одноклеточные организмы инфекциям и травмам? — Меротомия амеб и инфузорий. — Повреждения у Vaucheria. — Эпидемия, произведенная микросферою у амеб. — Внутриклеточное пищеварение у простейших. — Переваривание ими бактерий. — Эпидемия инфузорий. Болезнь ядра и ядрышка. — Деление инфицированных парамециумов и средство, благодаря которому они избавляются от паразитов. — Ацинеты — Хитридии.
Встречаются ли инфекционные болезни у одноклеточных организмов, столь распространенных в окружающих нас средах? Действуют ли и на них причины, вызывающие у нас воспаление?
Исследуем изменения, производимые такими причинами у низших организмов.
Даже самое незначительное повреждение неизбежно вызывает у человека и высших животных ряд типических воспалительных явлений. У одноклеточных существ все эти явления значительно упрощены. Если разрезать амебу пополам, то вдоль разреза не образуется даже раны, благодаря тому, что края заживают тотчас же после удаления лезвия (рис. 3, 4). В результате получаются две новые амебы. Та из них, в которой осталось первичное ядро, продолжает расти и представляет из себя нормальную особь; другая же, лишенная ядра, погибает через более или менее длинный срок *. Низшие протоплазматические существа, снабженные многими ядрами, как, например, Actynophrys **, могут быть разрезаны на несколько кусков; каждый из них регенерируется через очень короткое время, лишь бы в нем заключалась часть ядра ***.
У более дифференцированных инфузорий разрезывание острым инструментом производит настоящую рану, обнажающую внутреннюю плазму (рис. 5). Но края периферического слоя вскоре прикрывают рану и, выделяя новую кутикулу, зарубцовывают ее окончательно. Подобное заживление происходит как к кусках, снабженных ядром, так и в отрезках, лишенных его. Но в то время как у первых в короткое время (иногда меньше чем в 24 часа) совершается полная регенерация, вторые, напро-
* V. Bruno Hоfer. Experimentelle Untersuchungen uber den Einfluss des Kerns auf das Protoplasma. Jen. Zeitschr. f. Naturwissenschaft, т. XXIV, 1889, стр. 109, табл. IV, V.
** Очевидно, опечатка. Следует читать — Actinosphaerium, как и значится в названии цитируемой далее работы Брандта. — Ред.
*** V. Brandt, Ueber Actinosphaerium Eichhornii, 1887, стр. 30.
273
тив, постепенно атрофируются и всегда умирают. Бальбиани*, напечатавший весьма важную работу о «меротомии инфузорий», думает, что даже полное зарубцевание происходит исключительно в кусках, снабженных ядром.
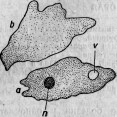
Рис. 3. Амеба, непосредственно вслед за разрезанием на две части: а — половина, заключающая ядро; b — половина, не заключающая ядра; v — бьющийся пузырек (по Бруно-Гоферу).
Рис. 4. Та же амеба через 5 минут после разрезания.
Ядро имеет очевидное влияние на выделение кутикулярной оболочки. У некоторых видов, как, например, Trachelius ovum, эктоплазма отдельных сегментов тотчас же после разрезывания покрывает рану; куски, снабженные ядром, регенерируются менее чем через пять часов.
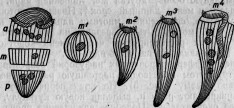
Рис. 5. Меротомия стентора: а — передний участок; т — средний; р — задний; m 1, т 2, т 3, т 4 — стадии регенерирующегося среднего участка (по Бальбиани).
Одноклеточные растения могут также подвергаться глубоким повреждениям, которые тем не менее не ведут их неизбежно к
* Recherches experimentales sur la merotomie des infusoires (Recueil zoologique suisse, т. V, 1888).
274
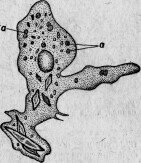
Рис. 6. Амеба в начале инфекции микросферами.
смерти. Таким образом, Ганштейн * наблюдал, что при разрезывании или раздавливании одноклеточной водоросли Vaucheria в ней отмирает только поврежденная часть; остальная же часть клетки вполне оправляется, выделив кутикулярную оболочку на пораненной области и образовав род секвестра. Следовательно, у низших организмов эти явления сводятся к более или менее полной и быстрой регенерации.
Помимо травмы, воспаление всего чаще вызывается инфекцией. Инфекционные болезни очень распространены между Protozoa и одноклеточными растениями, даже между их низшими представителями.
Мне удалось наблюдать у амеб эпидемию, производимую очень простым организмом в виде круглой клетки; он снабжен тонкой оболочкой и ядром и размножается делением.
Большая Amoeba с округленными псевдоподиями питается диатомеями. Но она содержит иногда, кроме них, небольшое количество вышеупомянутых круглых клеток (рис. 6), называемых мною Microsphaera. Протоплазматические движения и общий вид амебы остаются нормальными. Поэтому болезнь ее протекает незаметной по внешнему виду. Ближайшее наблюдение обнаруживает, однако, что только поглощенные диатомеи подвергаются пищеварительным изменениям, микросферы же, напротив, беспрепятственно размножаются делением внутри амебы. Мало-помалу последняя начинает выбрасывать диатомеи и становится все менее подвижной, обнаруживая этим свое болезненное состояние, протоплазма ее переполняется микросферами (рис. 7), которые в конце концов и убивают амебу.
Этот факт интересен тем, что показывает, как амеба, легко переваривающая даже двойчаток, может быть побеждена с виду
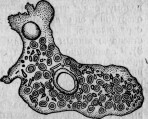
Рис. 7. Умирающая амеба, наполненная паразитическими микросферами.
* V. Frank. Die Krankheiten der Pflanzen, т. I, 1880, стр. 97.
275

Рис. 8. Амеба, живущая среди бацилл, часть которых поглощена ею.
совершенно слабыми существами, благодаря их способности противостоять ее пищеварительному действию.
Для объяснения этого явления приходится предположить, что паразит защищается, выделяя внутри амебы вещество, ограждающее его самого и ядовитое для нее.
Следовательно, инфекция развилась в амебе, несмотря на развитую у нее способность внутриклеточного пищеварения.
Ближайшее наблюдение мира простейших показывает, что пищеварительная функция играет существенную роль во взаимных отношениях этих животных. Многие корненожки и инфузории живут в средах, населенных массами других одноклеточных организмов, между прочим, бактериями. Эти последние, размножаясь крайне быстро, служат пищей многим простейшим. Таким образом, различные амебы поглощают бацилл, которые подвергаются в их теле целому ряду изменений. Так, не меняя своих очертаний, они приобретают способность легко окрашиваться везувином — способность, не свойственную живым бациллам вне организма (рис. 8). Совершенно подобные же изменения бацилл наблюдаются у сувоек (Vorticellae) и у других инфузорий, из чего очевидно, что они зависят от пищеварительного действия содержимого простейших. Это заключение вполне согласуется с наблюдением Б. Гофера* над пищеварением амеб. Он доказал, что чем большим изменениям подверглась пища внутри корненожки, тем лучше она окрашивается анилиновыми красками.
Часто приходится наблюдать, как жгутиковые монады поглощают нити Leptotrix, в несколько раз длиннее их самих (рис. 9), и кончают тем, что заключают их в свои пищеварительные вакуоли (рис. 10). Иногда можно проследить внутри инфузорий все изменения, происходящие с поглощенной бактерией.
Ле-Дантек ** сделал подобное наблюдение в моей лаборатории над поглощением стентором серной бактерии Thiocystis.
* Jen. Zeitschr., т. XXIV, 1889, стр. 109. ** Recherches sur la digestion intracellulaire, Lille, 1891, стр. 53.

Рис. 9. Монада, поглощающая нить лептотрикса.
276
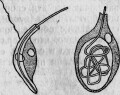
Рис. 10. Поглощение лептотрикса монадой.
Из приведенных фактов очевидно, что пищеварительное свойство протоплазмы простейших мешает распространению в них низших существ.
Эти последние только в исключительных случаях могут паразитировать в инфузориях и в корненожках.
Так как выше было уже упомянуто об одной из болезней корненожек, то я теперь прямо перейду к описанию эпидемии у инфузорий.
Давно уже было сделано наблюдение, что ядро некоторых видов инфузорий, а именно Paramaecium, заключает в себе очень тоненькие палочки. И. Мюллер, открывший их, думал, что это семенные тела. Другие исследователи, наблюдавшие их (Бальбиани и Бютчли), предположили, что эти палочки — паразитические бактерии. В действительности же эти организмы несомненно отличаются от бактерий и принадлежат к специальной группе, состоящей из нескольких видов. Два вида развиваются в ядре, совершенно выполняя его, а третий — исключительно в ядрышке. В своем вегетативном состоянии паразит этот является в виде удлиненных, веретенообразных клеток, размножающихся поперечным делением и дающих иногда почки. Достигнув зрелого состояния, паразиты превращаются в оригинальные споры, сходные по внешнему виду то с бациллами, то со спириллами* (рис. 11).
Несмотря на обилие паразитов в таких существенных органах, как ядро и ядрышко, зараженные инфузории не теряют способности делиться; но часто они гибнут вследствие истощения. Во время процесса деления часть паразитов зараженного Paramaecium выпадает из ядра и проникает в протоплазму животного, откуда выбрасывается, как всякое проглоченное, но не переваренное тело. Избавляясь таким образом при всяком последующем делении от некоторого количества своих паразитов, Paramaecium, поставленный в исключительно хорошие условия, как это было сделано в опытах г. Хавкина,
* См. работу г. Хавкина, сделанную в моей лаборатории и напечатанную в Annales de 1'Inst. Pasteur, т. IV, 1890, стр. 1888.
277
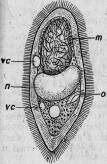
Рис. 11. Парамециум, ядрышко которого наполнено паразитами: о — рот; п — ядро; т — большое ядрышко; vc — бьющийся пузырек.
продолжает делиться, давая целый ряд все менее и менее зараженных поколений. Таким образом, в конце концов может получиться полное выздоровление. •
Г-ну Хавкину никогда не удавалось заразить Paramaecium спорами паразита. Он вводил инфузории в капиллярные трубки, заключающие эти споры. Будучи проглоченными и попав в пищеварительные вакуоли Paramaecium (рис. 12, 13), они выбрасывались подобно экскрементам.
Очевидно, что спора может прорасти в организме Paramaecium только в случае, если ей удалось избегнуть пищеварительного действия протоплазмы или быть выброшенной из нее. Для
прорастания она должна попасть прямо в ядро или ядрышко, лишенные этих функций.
Итак, мы видим на этом примере, так же как и из описанной болезни амеб, что микробу необходимо бороться против пищеварительных и выделительных функций протоплазмы.
Наибольшее количество инфекционных болезней Protozoae несомненно вызывается паразитизмом сосущих инфузорий, или ацинет, о которых я уже упоминал в первой главе.
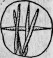
Рис. 13. Очень увеличенная вакуоля, заключающая споры.
Несмотря на тонкость своей кутикулы, эти паразиты отлично противостоят пищеварительному действию протоплазмы даже таких инфузорий (как, например, Stylonichiae), которые отличаются своей хищностью и легкостью переваривания добычи.
Как было уже упомянуто выше, молодые ацинеты прикрепляются к поверхности тела других инфузорий и, помощью активных движений, проникают в их эндоплазму.
Попав в центральную часть инфузории, паразиты значительно вырастают и делятся, производя большое количество молодых особей, до 50 и более. Некоторые из последних высвобождаются и после известного периода времени, проведенного вне организма, проникают в другую инфузорию.
Чтобы иметь возможность удержаться в протоплазме инфузорий, ацинеты должны оказывать какое-нибудь парализирующее влияние на их пищеварительную деятельность.
Очень вероятно, что эти паразиты выделяют какое-нибудь ядовитое вещество: часто наблюдали, что различные инфузории
278
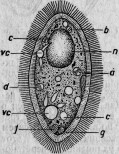
Рис. 12. Парамециум, поглотивший споры паразита: а, b, с, d, е, f, g — споры, окруженные вакуолями; п — ядро; vc — бьющийся пузырек.
были парализованы и умирали вследствие нападения свободных ацинет *.
Своим паразитизмом ацинеты вызывают дегенерацию ядра, которое распадается на круглые зерна. Часто, однако, болезнь, причиненная этими паразитами, не только не смертельна, но даже не мешает размножению инфузорий.
Для одноклеточных организмов гораздо опаснее инфекционные болезни, производимые низшими грибами из группы Chitridiae. Впрочем, эти паразиты внедряются обыкновенно в простейших, питающихся не твердой пищей, а исключительно эндосмотически. На инфузорий же, способных к внутриклеточному пищеварению, они нападают только во время их покоящегося состояния — кисты, когда пищеварение прекращено.
Внутриклеточные паразитические Chitridiae, проникнув в инфузорию,

Рис. 14. Зеленая евглена, заключающая хитридия.
Рис. 15. Евглена, наполненная зооспорами хитридия.
принимают вид круглой неподвижной клетки. В этом состоянии они питаются на счет содержимого инфузории, которая в конце концов умирает. Сами же они обращаются в зооспоры.
Возьмем для примера Chitridium, паразитирующего в зеленой евглене и открытого Клебсом. ** Евглены водятся в стоячих водах. Между ними встречаются особи, с виду совершенно здоровые, но заключающие круглое тело, снабженное ядрами и очень тонкой оболочкой (рис. 14). Это тело постоянно вырастает и делится на большое количество мелких клеток (рис. 15), превращающихся в конические зооспоры. Они пробуравливают евглену и выходят в окружающую воду.
Во время развития паразита в самой евглене проявляются ясные признаки болезни: ее зеленые хроматофоры быстро резорбируются, и она принимает в высшей степени анемичный вид. Ее содержимое подвергается в то же время пигментному перерождению, выраженному в образовании отдельных бурых зерен, число которых все более и более увеличивается. Евглена обыкновенно погибает в то время, когда паразит достиг стадии зооспоры. Инфекция, производимая хитридием, наблюдается у евглены только в ее подвижном состоянии, так что, очевидно,
* См. Бютчли. Protozoae, в Bronn's Klassenu. Ordnungen des Thierreichs, т. III, 1889, стр. 1823, 1842.
** Untersuchungen aus d. botan. Instit. in Tubingen, т. I, 1883. Также Хавкин. Annales des sciences naturelles. Zoologie, 1886, стр. 330, 336 и т. д.
279
киста, которой она окружена в покоящемся состоянии, служит ей защитой. Однако она оказывается недостаточной против нападения другого представителя хитридий, Polyphagus euglenae, который легко развивается в ней.
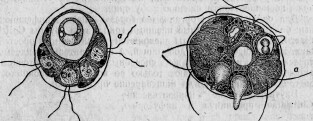
Рис. 16. Пандорина, одна из клеток которой заражена ольпидиумом.
Рис. 17. Другая пандорина с пятью зараженными клетками.
Колониальные жгутиковые точно так же подвергаются инфекции хитридий. Один из представителей Volvocineae — Pandorina morum — часто бывает заражена паразитом из рода Olpidium. Последний вызывает со стороны Pandorina выделение жидкости, скопляющейся в вакуоли (рис. 16). Прозрачная маленькая паразитическая клетка растет на счет пандорины и наполняется жировыми зернами и вакуолями. Вскоре паразит образует конический отросток, пробуравливающий оболочку пандорины (рис. 17, 18). Через этого отросток проходят зооспоры, развившиеся в паразите. В других случаях Olpidium не дает
зооспор, а прямо закистируется, выделив толстую оболочку.
Зараженная клетка пандорины, претерпевая, как и евглена, пигментное перерождение, распадается и в конце концов всегда умирает. Что же касается других особей колонии, даже ближайших соседей погибшей клетки, то они остаются совершенно невредимыми: сохраняют полную подвижность, сократительность бьющихся пузырьков и продолжают совершенно нормально размножаться делением (рис. 18). Таким образом, болезнь и смерть одной или даже

Рис. 18. Зараженная часть пандорины; а — зооспорангий паразита.
280
большинства из шестнадцати членов колонии нисколько не отражается на особях, избегнувших паразита.
Этот очерк искусственных повреждений и инфекционных болезней одноклеточных организмов прежде всего обнаруживает недостаточность современных сведений по этому вопросу. Но он дает нам в то же время основание для оценки общего характера изложенных фактов. В явлениях, следующих за повреждением, нас всего более поражает способность полной регенерации. Мы видели, что отрезанный сегмент может через очень краткий срок, несколько часов или даже минут, вернуться к своему нормальному состоянию.
Из фактов, изложенных в первой главе, можно смело вывести общее положение, что отношения между простейшими и инфицирующими организмами сводятся к активной борьбе. Паразиты часто не что иное, как хищники, которые вследствие своих малых размеров не могут прямо нападать на добычу, а внедряются в нее, питаясь на ее счет.
Эта тесная связь между паразитизмом и хищничеством проявляется не у одних ацинет, но и у других паразитичных инфузорий (Vampyrellae и др,) 158. При инфекции борьба усложняется и становится только менее непосредственной. Нападение паразита в этих случаях сводится к выделению ядовитых или растворяющих веществ; защищается же он, парализируя способность своей добычи переваривать и извергать. Последняя, в свою очередь, борется, стараясь переварить или извергнуть паразита, и защищается выделениями, которыми себя окружает.
Хотя все эти явления не входят в понятие борьбы за существование в строго дарвиновском смысле слова (т. е. в понятие о конкуренции между индивидуумами одного вида с целью переживания и производства лучше приспособленного потомства). однако они сводятся к более или менее непосредственной борьбе между представителями различных групп организмов.
Важная роль в этой борьбе принадлежит внутриклеточному пищеварению, столь распространенному у корненожек и инфузорий, и присущему даже простейшим, питающимся осмотическим путем.
ЛЕКЦИЯ ТРЕТЬЯ
Пласмодий. Уколы стеклянной трубкой. — Прижигание нагретой палочкой. — Химические возбуждения. — Трофотропизм. — Химиотаксия. — Приучение пласмодия к различным веществам. — Роль отрицательной химиотаксии. — Отталкивание от бактерий. — Переваривание бактерий пласмодием. — Чувствительность пласмодия. — Неподвижные растения лишены настоящего внутриклеточного пищеварения. — Некроз и регенерация. — Опыты Вальденбурга. — Роль оболочки. — Исследования де-Бари относительно Peziza sclerotiorum. — Опухоли растений
Перейдем к рассмотрению патологических явлений у многоклеточных организмов. Мы остановимся при этом сначала на очень важной группе, интересной главным образом простотой своего строения. Я имею в виду миксомицетов, соединяющих в себе животные и растительные признаки. Одна из стадий их развития — пласмодий — представляет наибольшую массу протоплазмы, существующую в природе.
Как известно, пласмодий есть не что иное, как амебоидное состояние миксомицетов, образованное скоплением большого количества зооспор в одну протоплазматическую массу со множеством ядер.
Разветвляясь в различные направления, пласмодий может передвигаться по сухим листьям, дереву и другим предметам, на которых он находится. Края его эктоплазмы двигаются амебоидно; в эндоплазме же наблюдаются быстрые течения, подобные потокам лавы.
Пласмодий легко поглощает твердые тела, находящиеся на его пути. Он переваривает их с помощью пептического фермента и кислоты, выделяемой вокруг пищи *.
Все же остатки, так же как и непереваренные тела, выбрасываются им.
В определенный период своей жизни пласмодий дает спорангии. Они имеют всего чаще форму маленьких плодов, и содержимое их распадается на множество спор, снабженных твердой оболочкой.
Пласмодий достигает иногда очень значительных размеров — иногда до фута и более в длину. Благодаря этому он представляет громадные преимущества для изучения протоплазмы вообще и ее патологических явлений в частности.
* Пептический фермент был открыт Крукенбергом: Untersuchungen aus d. physiol. Inst. d. Univ. Heidelberg, т. II, 1878, стр. 273. Относительно кислоты пласмодия см. Annales de 1'Inst. Pasteur, 1889, стр. 25.
282
Начнем с исследования травматического влияния на пласмодий миксомицета Physarum. Введем в его протоплазму твердое тело, например, маленькую стеклянную трубку. Во время укола конец трубки разрывает пласмодий, часть которого вытекает в окружающую жидкость. Но большая масса протоплазмы остается совершенно невредимой и обволакивает трубку (рис. 19), как всякое постороннее тело, годное для питания. Через более или менее короткое время пласмодий извергает ее, подобно всяким негодным остаткам.
Мы можем прибегнуть к другому средству для раздражения пласмодия. Возьмем для этого желтый Physarum, растянутый на предметном стекле, и прижжем центральную часть его рас-
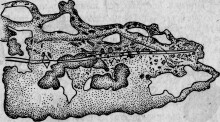
Рис. 19. Часть пласмодия, поглощающая стеклянную трубку.
каленной стеклянной палочкой. Мы произведем таким образом термическое раздражение вместо механического, как в первом случае. Тотчас после обжога центральная часть пласмодия отмирает. Вид ее резко отличается от окружающих периферических, живых частей. Последние сохраняют неподвижность и оставляют умерший участок нетронутым. Однако через несколько часов пласмодий выходит из этого пассивного состояния и, удаляясь, покидает омертвелую часть.
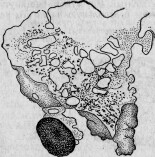
Еще более резкое действие имеют химические возбудители. Положивши кусочек ляписа на край пласмодия Physarum, растянутого на предметном стекле, обмоем тотчас поврежденную часть 1 % раствором поваренной соли (для осаждения излишка ляписа); тогда мы увидим, что прижженная часть отмерла и отделилась от остального пласмодия (рис. 20). Реакция последнего выражается резким изменением направления его движений. Опыт был намеренно произведен в той части пласмодия, по направлению к которой шло течение эндоплазмы. Тотчас после прикосновения ляписа течение эндоплазмы приняло диаметрально противоположное направление (рис 21). Через час после
283
начала опыта пласмодии уже удалился со своего первоначального места, оставив на нем омертвелый участок.
Общая черта двух последних опытов заключается в удалении пласмодия от части, поврежденной термическим или химическим раздражителем. Только в последнем случае удаление происходит гораздо быстрее, что объясняется влиянием ляписа.
Итак, мы видим, что раздражители вызывают в пласмодии то явления, сходные с поглощением твердой пищи, то явления более или менее резко выраженного отталкивания. Желая вызвать реакцию, соответствующую воспалению у высших животных, мы натолкнулись на явления притяжения и отталки-
вания, столь распространенные в жизни пласмодиев и низших существ вообще.
Уже в 1884 г. Сталь * открыл, что настой сухих листьев, служащий питательной средой для стольких миксомицет, притягивает пласмодии. Напротив, растворы солей, сахара и некоторых других веществ действуют обратно. Они отталкивают пласмодии, заставляя их удаляться на более или менее значительное расстояние.
Сталь, проводя аналогию между этими явлениями и питанием, случаи притяжения называет положительным, а отталкивания — отрицательным трофотропизмом. С другой стороны, Пфеффер ** доказал, что женские половые органы некоторых споровых (папоротников, мхов и селагинель) притягивают семенные тела. Так как цель этого притяжения не есть питание, то он обозначил все проявления чувствительности к химическим
* Botanische Zeitung, 1884, № 10—12. ** Untersuchungen aus d. Botan. Inst. Tubingen, т. I, стр. 363.
284
Рис. 20. Пласмодий, прижженный ляписом.
Рис. 21. Тот же пласмодий через 50 минут после стадии, изображенной на рисунке 20.

агентам общим именем химиотаксии (положительной или отрицательной). Это название стало вскоре общепринятым *.
Мы видели, что эти явления чувствительности играют несомненную роль в патологических процессах. Поэтому следует несколько ближе познакомиться с ними.
Химиотаксические свойства присущи не только миксомицетам и семенным телам упомянутых споровых. Они встречаются также у бактерий, у жгутиковых инфузорий, у Volvocineae ** и у зооспор грибов, например, Saprolegniae ***. Такое распространение этого явления доказывает общность его характера. Несомненно, что положительная химиотаксия руководит организмами в поисках питательных веществ; она же направляет их к полезным в других отношениях телам, как, например, в случаях притяжения семенных тел яйцевой клеткой.
Отрицательная химиотаксия служит, наоборот, средством защиты от вредных влияний. Это правило, верное вообще, может не быть применимо к каждому частному случаю. Так, Пфеффер **** наблюдал смерть спирилл и Bodo, вызванную их проникновением в слишком насыщенный раствор сахара или глицерина, к которым были примешаны притягательные вещества.
Аналогия этих явлений с чувствительностью человека и высших животных кидается в глаза. Она подтверждается, между прочим, тем, что закон Вебера, установленный для чувствительности человека, распространяется и на химиотаксию низших существ.
Так, для того чтобы бактерия (Bacterium termo) или семенное тело папоротника ***** заметили разницу в составе данной среды, надо, чтобы она изменилась в определенной пропорции. Например, для того чтобы Bacterium termo, находящийся в определенном пептонном растворе, направился в более насыщенный, надо, чтобы концентрация последнего была в пять раз сильнее первого. Пфеффер, установивший эти отношения, применил к химиотаксии одноклеточных закон, управляющий чувствительностью человека. Закон этот заключается в том, что возбуждение растет в геометрической, чувствительность же в арифметической прогрессии. Реакция, следовательно, пропорциональна логарифму возбуждения.
Однако, несмотря на применимость этого закона как к химиотаксии одноклеточных, так и к чувствительности человека,
* В настоящее время обычно говорят о хемотаксисе или хемотропизме. — Ред.
** Там же, т. II, 1888, стр. 582. *** Botanische Zeitung, 1890, № 7—11.
**** Untersuchungen aus d. Botan. Inst. Tubingen, т. II, стр. 627. ***** Организмы, над которыми Пфеффер сделал свои замечательные наблюдения.
285
разница между этими явлениями очень велика в количественном отношении.
Так, человек чувствует изменение веса на 1/3, температуры на 1/30, света на 1/100. Семенные же тела папоротника замечают изменение концентрации среды только если она увеличилась в 29 раз, a Bacterium termo — в 4 раза (Пфеффер, 1. с., II, стр. 637).
Чтобы составить себе понятие о химиотаксической чувствительности пласмодия (Dydimium farinaceum), я поместил несколько образчиков его в растворы солянокислого хинина различной концентрации: 0,1; 0,01; 0,05; 0,005 и 0,0005%. Оказалось, что пласмодии не только приближались, но и погружали свои отростки в два последних, более слабых раствора.
Напротив, три первых, более сильных раствора вызывали в них явственную отрицательную химиотаксию. Следовательно, пласмодии чувствителен к изменению концентрации хинного раствора от 0,05 до 0,005%.
Очень важным свойством как пласмодия, так и других низших организмов является постепенное их приучение к таким растворам, которые не переносились ими вначале.
Сталь первый сделал наблюдение, что пласмодии Fuligo сначала избегает 2% раствора поваренной соли. Но, за недостатком пресной воды, он постепенно приспособляется к соленой и погружает в нее свои отростки. Это служит нам примером того, что под влиянием неуловимых изменений, происшедших в протоплазме, отрицательная химиотаксия может обратиться в положительную.
Так как это явление имеет громадное общее значение, то я постарался воочию убедиться в нем.
Для этого я поместил пласмодии Physarum, растянутый на предметном стекле, в сосуде с 0,5% раствором хлористого натрия. Пласмодии тотчас же обнаружил отрицательную химиотаксию и удалился от поверхности жидкости. Тогда я перенес его в 0,25% раствор той же соли. Сначала он удалился, но через несколько часов приблизился к этой жидкости и погрузил в нее свой край. Ввиду такого приспособления, я вновь перенес пласмодии в 0,5% соляной раствор. Сначала он удалился, но приблизительно через 12 часов спустился к поверхности жидкости, не погружая, однако, в нее своих отростков.
Итак, отрицательная химиотаксия служит для пласмодия средством избежания вредных влияний. Мы видели, что он удаляется от прижигающих его тел, как ляпис, и даже собственных омертвелых частей, как в случае ожога раскаленной стеклянной палочкой. Это же свойство, по всей вероятности, предохраняет пласмодии от нападения других организмов, а именно от патогенных микробов.
Сталь утверждает, что у пласмодиев никогда не бывает паразитов. Он объясняет этот факт как подвижностью пласмодиев,
286
так и способностью их отбрасывать инородные тела. Способность эта связана у них с внутриклеточным пищеварением. Хотя никогда еще не удавалось непосредственно наблюдать выбрасывания пласмодием паразитических организмов, тем не менее предположение это не имеет ничего невероятного. Так, Пфеффер* видел, как пласмодий Chondrioderma выбрасывает живых Pandorinae и Diatomeae. Предположение Сталя подтверждается еще тем, что парамециум несомненно выбрасывает паразитические споры.
Для выяснения значения подвижности пласмодия я сделал следующий опыт. Пласмодий Physarum был помещен на предметное стекло в равном расстоянии от двух сосудов. Один из них был наполнен старым настоем из сухих листьев с бактериями и другими простейшими. Другой же заключал тот же раствор, но только предварительно несколько раз профильтрованный.
Края пласмодия были соединены с содержимым каждого сосуда посредством полосок из промокательной бумаги. Очень скоро пласмодий направился на бумажную полоску, пропитанную фильтрованной жидкостью.
Точно такие же результаты дал другой, несколько видоизмененный опыт. Из этого видно, что пласмодий предпочитает растворы, очищенные от микробов.
Чтобы выяснить степень этого предпочтения, я повторил тот же опыт, заменив только фильтрованную жидкость свежим, следовательно, бесцветным, настоем сухих листьев в холодной воде. На этот раз пласмодий направился к старому настою, несмотря на присутствие в нем микробов.
Следовательно, отталкивание его от низших организмов имеет свои пределы. Это вполне согласуется с фактом, что миксомицеты в амебоидном состоянии поглощают микробов. Сэвиль Кент наблюдал амебоидные зооспоры Physarum tussilaginis, наполненные бактериями. Затем Листер ** сделал очень интересные наблюдения над поглощением бактерий зооспорами различных миксомицетов. Бактерии захватываются псевдоподиями, втягиваются в плазму и препровождаются в пищеварительные вакуоли. Здесь они становятся все менее и менее явственными, точно растворяются. Наблюдали, как зооспора Chondrioderma difforme вполне переварила в течение l1/2 часов двух больших бактерий.
Пищеварительная и выделительная функции пласмодия, связанные с отрицательной химиотаксией, могут быть очень полезны в реакции против различных вредных раздражителей.
* Ueber Aufnahme und Ausgabe ungeloster Korper. Abhandlungen d. mathem.-physikalischen Classe der K. Sachs. Gesellsch. d. Wiss., т. XVI, 1890. стр. 161.
** Journ. of the Linnean Society, т. XXV, Botany, 1890, стр. 435.
287
Но пласмодии, так же как и многие другие низшие организмы, одарены чувствительностью, помимо химиотаксической. Например, они избегают солнечного света и сильно притягиваются сыростью. Они обнаруживают, следовательно, род гидротропизма. Перед плодоношением этот положительный гидротропизм обращается в отрицательный, и пласмодий ищет сухих мест (Сталь). Кроме того, он обладает осязательной чувствительностью.
Мы видели, что пласмодии избегают вредных влияний передвижением или же избавляются от них путем выделения и пищеварения. Что же касается многоклеточных растений, не одаренных ни подвижностью, ни способностью переваривать и выделять инородные тела, то они должны другим путем реагировать против вредных влияний.
Пласмодии реагирует против введенной в него занозы точно так же, как и против всякого поглощенного им непереваримого тела, т. е. выбрасывает ее. Но та же заноза, введенная в ткань высшего растения, неизбежно производит смертельное повреждение затронутой клетки. Потеря вещества, происшедшая вследствие этого, восстановляется быстрым размножением соседних клеток. Они образуют то древесную ткань, то настоящий рубец из различных тканей *. В обоих случаях мы имеем дело с активным новообразованием клеток на поврежденном месте. Вальденбург** изучал эти явления с точки зрения их аналогии с воспалением у высших животных. Он находит сходство между характеристическими признаками рассматриваемых процессов.
«Воспаление, — говорит он, — встречается, следовательно, и у растений. Но под словом «воспаление» надо понимать только повреждение и вызванную им опухоль, без вмешательства сосудов и нервов».
Воспаление, с этой точки зрения, есть исключительно раздражение тканей (опухоль, разрастание), сопровождаемое приливом крови (стр. 344).
Часто приводят процесс восстановления у растений в подтверждение теории «притяжения» при воспалении. Особенно же часто приводят его в пользу теории Вирхова об усиленной деятельности питания и размножения воспаленных тканей. Но, к несчастию, при этом совершенно игнорируют промежуточные формы между растениями и высшими животными. Вследствие этого и были упущены явления, именно наиболее характерные для воспаления.
Новые клетки, развившиеся на поврежденных частях растения, часто выделяют оболочку еще более толстую и плотную, чем первоначальная. Оболочка есть настоящий орган защиты
* V. Frank. Die Krankheiten der Pflanzen, Breslau, 1880, стр. 1—95. ** Archiv fur pathol. Anat. u. Physiol. Virchow's Archiv, т. XXVI, 1863, стр. 145 и 322, табл. V
288
растения. Мы это видим из примера евглены (приведенного в предыдущей главе), для которой нападение хитридия опасно только в подвижном состоянии; в покоящемся же киста служит ей защитой. Растительная оболочка слишком упорна для многих микробов, особенно для тех, которые не в состоянии активно проникнуть внутрь клетки. Этим, вероятно, объясняется редкость бактериальных инфекций у растений. Последние, напротив, очень часто подвержены нападению грибов, обладающих значительной силой прорастания. Кроме того, некоторые из них выделяют фермент, растворяющий клетчатку растительной оболочки. Войдя внутрь клетки, паразитический гриб беспрепятственно поглощает ее содержимое. Зараженные клетки погибают, другие же, оставшиеся живыми, усиленно размножаются. Это приводит к гипертрофии частей и иногда даже целого заболевшего организма (например, у Euphorbia cyparissias под влиянием эцидия Uromyces pisi). Часто паразит вызывает образование специальных опухолей, или чернильных орешков, развивающихся под влиянием грибов точно так же, как и от действия животных организмов. Как при заживлении ран, так и при инфекции* у растений наступают восстановительные явления вследствие усиленного размножения клеток, не поврежденных непосредственно, не представляя процессов, сходных с воспалительными. Чтобы познакомиться с этими последними, надо перейти к обзору представителей животного царства.
* Лучше всего изученный пример инфекции у растений представляет Peziza sclerotiorum, о которой де-Бари сделал классическую работу (Botan. Zeit., 1886). Гриб этот растет на поверхности растения и пускает нити, с помощью которых проникает внутрь его (Peziza scl. заражает многие растения). В начале своей паразитической жизни нити гриба выделяют щавелевую кислоту и фермент, растворяющий клетчатку.
Паразит питается на счет сока клеток, убитых его выделениями, и проникает своим мицелием в промежутки между клетками, только в редких случаях входя внутрь их. Де-Бари наблюдал, что Peziza легко проникает в молодые растения, но встречает непобедимые препятствия для проникновения внутрь более взрослых. Очень вероятно, что иммунитет этих последних происходит вследствие невозможности для гриба растворить клетчатку старых клеток. Контрольные опыты показали, что, действительно, выделения грибов легко переваривали оболочки молодых клеток и оставляли их неизменными у более взрослых клеток тех же растений. Очевидно, что сопротивление растительной клетки основано главным образом на свойствах ее оболочки. Для произведения инфекции паразит должен прежде всего пробуравить или растворить последнюю [т. е. оболочку].
ЛЕКЦИЯ ЧЕТВЕРТАЯ
Переход одноклеточных к многоклеточным. — Беглый очерк теории Phagocytella. — Protospongia. — Губки; их организация. — Три зародышевых пласта. — Питание губок. — Внутриклеточное пищеварение. — Искусственное деление. — Введение острых тел. — Утилизация посторонних тел для скелета. — Судьба организмов, проникших в тело губок. — Роль эктодермы в защите. — Сравнение с миксомицетами. — Сравнение с воспалением позвоночных
Переходя к животному царству, мы должны прежде всего отметить тот факт, что, к сожалению, до сих пор не знают, каким образом многоклеточные животные произошли из простейших. Пропасть, лежащая между наиболее совершенными представителями этих последних и наиболее низшими многоклеточными, слишком велика и может быть пополнена только гипотезами, основанными на эмбриологическом изучении различных животных. Оставляя в стороне некоторые группы паразитов, утративших многие из своих первоначальных признаков (Dycyemidae, Ortonectidae), обратимся к простейшим многоклеточным, как, например, губки. Они состоят уже из целой суммы органов, образованных тремя пластами — эктодермой, мезодермой и энтодермой. Чтобы получить понятие об еще более простом состоянии животных, надо обратиться к зародышам губок и других низших животных, как, например, медуз и их родичей. Здесь можно легко найти стадии, состоящее из двух зародышевых пластов. Один из них представляет наружную оболочку личинки, тогда как другой образует группу внутренних клеток, представляющих различные особенности. Эти клетки то образуют плотное скопление, род паренхимы, составленной из амебовидных элементов, то они равномерно расположены в один эпителиальный слой, выстилающий пищеварительную полость. Много спорили о том, которая из этих двух форм должна быть рассматриваема как первичная. Я думаю, что отсутствие пищеварительной полости, неправильная форма клеток и целый ряд других доказательств из сравнительной эмбриологии многих низших животных (о которых я не могу говорить в этом патологическом очерке) позволяют рассматривать паренхиматическую стадию как простейшую. Этой стадии я дал название phagocytella *, вследствие способности клеток нижнего слоя захватывать различные твердые тела и особенно вследствие того,
* См. изложение теории phagocytella в Embryologische Studien an Medusen, Wien, 1886 159.
290
что этот слой производит пищеварительные клетки готового организма. Сначала он производит энтодерму, выстилающую пищеварительный канал с его придатками, а затем всю мезодерму или только ее часть, в которой также заключается большое число пищеварительных клеток, или фагоцитов.
Стадия phagocytella может легко превратиться в стадию гаструла*, состоящую из двух эпителиальных слоев, из которых один представляет стенку первоначального пищеварительного канала, открывающегося особым отверстием, или бластопором. Эта гаструла объясняет много различных стадий других многоклеточных.
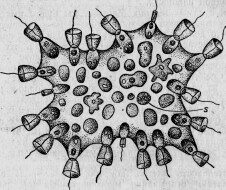
Рис. 22. Protospongia Haeckeli (по Савиль Кенту).
Организм этих последних в своей наиболее примитивной форме, именно в виде phagocytella, представляет аналогию с некоторыми колониальными формами простейших, у которых колонии составлены из двух различных видов клеток: жгутиковые индивидуумы, образующие род наружного слоя (рис. 22), и амебовидные — внутреннюю часть колонии. Первые соответствуют эктодерме, составленной частью из жгутиковых клеток; вторые представляют как бы внутреннюю паренхиму, состоящую из амебовидных и в то же время фагоцитных клеток. В этих инфузориях, названных Protospongia (это название дано Савилем Кентом**, который их открыл), два слоя не представляются еще ясно разграниченными, так как составляющие их индивидуумы легко могут превращаться друг в друга 160
* См. теорию Gastrea: Наесkе1. Gastrea-Theorie, Jena, 1874. ** The Manual of infusoria, 1880—1882.
291
Таким образом, является возможность воспроизвести связь между простейшими и многоклеточными животными через жгутиковых колониальных инфузорий, с одной стороны, и посредством организмов, похожих на phagocytella, — с другой.
Я бы никогда не остановился на этих гипотезах в курсе сравнительной патологии воспаления, если бы изложение их не представляло нам случая познакомиться с общим значением амебовидных клеток, способных захватывать твердые тела. Найдя их в различных классах простейших, мы вновь встречаемся с ними в самых низших формах многоклеточных. Участие амебовидных клеток в воспалении позвоночных есть общепризнанный факт громадной важности.
Начиная с самых низших Metazoa, мы будем иметь дело с этими клетками. Губки имеют настолько мало развитую организацию, что долгое время на них смотрели, как на колонии простейших, составленные, подобно Protospongia, из жгутиковых амебовидных индивидуумов. Только позднее установили известное родство губок с полипами и с другими близким им формами (Coelenterata). С тех пор вполне убедились, что губки состоят из трех характерных слоев. Поверхностный слой, или эктодерма, покрывает все тело плоскими эпителиальными клетками. Они разграничены между собой контурами, которые становятся очень заметными после действия на них азотнокислым серебром. Сами клетки заметно сократимы; это особенно легко наблюдается на свободных краях молодых индивидуумов, где можно заметить амебовидные удлинения эктодермических элементов.
Сократительность клеток, конечно, играет роль в замечательном явлении открывания многочисленных пор, разбросанных на поверхности всей губки между каждыми двумя или несколькими плоскими клетками. Эти поры открываются, чтобы впустить струю воды и мельчайшие тела, которые находятся в ней во взвешенном состоянии.
Жидкость проникает сначала в систему вводящих каналов, также выстланных плоским эпителием, происхождение которого еще точно не определено. Затем она проходит в каналы или в расширения, называемые «мерцательными корзинками». Последние покрыты цилиндрическим эпителием, клетки которого снабжены одним большим жгутиком. Эти клетки, обнаруживающие поразительное сходство с многими жгутиковыми инфузориями, принадлежат эктодерме * и представляют настоящих фагоцитов.
Большое количество маленьких зернышек, принесенных потоком, привлекается этими эктодермическими * клетками и поглощается ими.
* Повидимому, опечатка. Следует читать «энтодерме», «энтодермическими». — Ред.
292
Но, кроме жгутиковых фагоцитов эктодермического * происхождения, в губках есть еще и другие, являющиеся в форме подвижных клеток. Они представляют настоящих маленьких амеб, расположенных между эктодермой и цилиндрическим эпителием, и принадлежат к мезодерме. Хотя еще недостаточно известен способ, посредством которого посторонние тела, находясь внутри губки, проникают в мезодерму, однако уже доказано, что эти тела в большом количестве поглощаются самими мезодермическими клетками. Если прибавить к воде, в которой живут губки, какое-нибудь красящее вещество, например кармин, индиго или сепию, то скоро можно заметить, что зерна краски поглощаются эктодермическими ** клетками, а также и амебовидными фагоцитами мезодермы.
У некоторых видов губок (например, у многих известковых) мезодермические клетки немногочисленные и играют второстепенную роль в захватывании посторонних тел; у других (особенно у кремневых) мезодерма преобладает, и ее фагоциты поглощают большое количество введенных тел. Есть некоторые губки, как, например, Siphonochalina coriacea, у которых исключительно мезодермические клетки захватывают посторонние тела. Цилиндрические же клетки эктодермы*** служат только для поддержания постоянного протекания жидкости через организм губки.
Фагоциты обоих слоев могут выбрасывать нерастворимые вещества. Эти последние скопляются в больших выводящих каналах и выталкиваются наружу помощью крупных отверстий в форме кратеров. Стенки их, по наблюдению некоторых авторов, снабжены мускульными волокнами 161.
Факт, интересующий нас более всего, состоит в том, что мезодермические фагоциты способны не только захватывать посторонние тела и выбрасывать их нерастворимые остатки, но также могут и переваривать различные вещества, приходящие к ним из окружающей среды. Либеркюн **** уже давно наблюдал переваривание инфузорий, проникших в амебовидные клетки пресноводных губок. Он проводил аналогию этого явления с перевариванием инфузорий протоплазмой корненожек и других простейших. Этот факт был подтвержден и другими наблюдателями. Я сам мог констатировать ***** растворение Охуtricha, Glaucoma и Actinophrys среди мезодермических фагоцитов молодых Spongillae. Посторонние тела, проглоченные этими простейшими, были затем также захвачены мезодермическими фагоцитами. Евглены, увлеченные потоком в тело Spongillae, таким же образом окружаются фагоцитами мезодермы. Прото-
* Следует читать «энтодермического». — Ред.
** Энтодермическими. — Ред.
*** Энтодермы. — Ред.
**** Muller's Archiv fur Anatomie und Physiologie, 1857, стр. 385.
***** Zeitschr, f. wiss. Zool. т. XXXII, 1879, стр. 371.
298
плазма этих захваченных жгутиковых переваривается; парамиловые же и хлорофильные зерна остаются нетронутыми неопределенно долгое время.
Мезодермические клетки молодых Spongillae, происшедших из геммулы, захватывают посторонние тела даже в то время, когда эктодерма еще совсем не образовалась. Молодая губка состоит тогда только из слоя плоских эктодермических клеток и неправильной массы клеток мезодермы. Многие из последних скоро начинают выделять иглы скелета. Зерна кармина, пла-
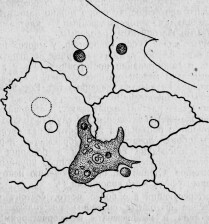
Рис. 23. Мезодермический фагоцит молодой Spongilla, окруженный несколькими эктодермическими клетками.
вающие в воде, где живут губки, проникают внутрь этих последних и захватываются амебовидными фагоцитами мезодермы. При проникновении кармина во внутренность губок не замечается никакого видимого повреждения их стенок (рис. 23). Так как важная роль амебовидных клеток мезодермы у губок при захватывании и переваривании посторонних тел не может быть оспариваема, то я постарался вникнуть в условия, при которых совершаются эти явления. На основании того, что многие простейшие и миксомицеты представляют собой пример внутриклеточного пищеварения, выделяя при этом вокруг захваченных тел некоторое количество кислоты, я впускал в воду, где жили молодые губки, зерна синего лакмуса. Эти последние очень скоро захватывались губками и особенно фагоцитами
294
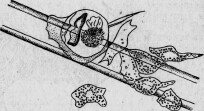
Рис. 24. Стеклянная трубка, окруженная мезодермическими фагоцитами Spongilla.
мезодермы. Но, несмотря на долгое пребывание в этих клетках, лакмус не изменял своего цвета. Это доказывает, что пищеварение губок не совершается в кислой среде. Факт этот вполне согласуется с открытием Крукенберга *, нашедшего трипсический фермент в глицеринной вытяжке многих губок.
Легко наблюдать то, что происходит в организме губки, когда в нее вводят какое-нибудь острое тело, например тоненькую стеклянную трубочку или иглу асбеста. Предмет проникает в мезодермическую массу, где и находится в соседстве с амебовидными клетками. Эти клетки окружают его вполне или только отчасти, реагируя, как будто бы это постороннее тело представляло собой питательную массу, но больших размеров, чем обыкновенно (рис. 24). Иногда клетки вовсе или почти совсем не собираются вокруг введенного тела.
Это показывает, что оно не возбудило достаточной реакции. Иногда же какой-нибудь незначительный предмет, например растительная нить, привлекает значительное количество фагоцитов, которые ее окружают, сливаясь в маленькие пласмодии (рис. 25). У многих губок зерна песка и другие твердые тела, попавшие в них случайно, окружаются массой спонгина, выделяемого мезодермическими клетками. В подобных случаях эти посторонние тела утилизируются губкой, увеличивая прочность ее скелета.

Рис. 25. Растительная нить, окруженная фагоцитами Spongilla.
* Grundzuge einer vergleichenden Physiologie der Verdauung. Heidelberg, 1882, стр. 52.
295
Как было сказано выше, мезодермические клетки могут окружать и переваривать также и живые организмы, проникшие во внутренность губки. Более прочные организмы не поддаются легко этому разрушительному действию фагоцитов и могут пребывать более или менее долгое время без всякого изменения в

Рис. 26. Leptotrix, окруженный фагоцитами Spongilla.
теле губки. Так, я наблюдал пучки Leptotrix среди мезодермы молодых губок. Фагоциты, соединенные в пласмодии, окружали этих бактерий, причем последние оставались совсем неизмененными (рис. 26). Келлер нашел внутри некоторых губок (Hircinia echinata и Ceraochalina gibbosa) яйца кольчатых червей
и ракообразных. Эти яйца свободно развиваются среди мезодермы; они окружены скоплением амебовидных клеток, образующих настоящую фолликулу (рис. 27).
Из вышеизложенных фактов следует, что вообще все посторонние тела, попавшие как-нибудь в паренхиму губок, возбуждают деятельность мезодермических фагоцитов. Эти последние захватывают тогда внутрь себя эти тела или окружают их, соединяясь или сливаясь по нескольку вместе.
Если захваченные посторонние организмы легко перевариваемы, то и подвергаются этому процессу; если же, наоборот, они непереваримы, то остаются в губке, окруженные клетками, и представляют собою род «нахлебников» 1б2.
296
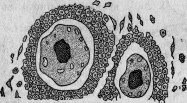
Рис. 27. Яйца ракообразного, окруженные фагоцитами Ceraospongilla (по Келлеру).
Явление это очень распространено среди губок. Эти мягкие животные, проникновение в которых не затруднительно, становятся удобным убежищем для многих водных существ. Губки доставляют им пищу, поддерживая постоянный приток воды с различными плавающими в ней телами. Известно, например, большое число нахлебников губок, начиная с водорослей (Zoochlorellae, Zooxantellae), живущих внутри мезодермических клеток, и кончая полипами (Stephanoscyphus), кольчатыми и ракообразными, населяющими каналы и паренхиму губок. До сих пор еще неизвестны настоящие паразиты губок, а вследствие этого не знают еще и их инфекционных болезней. Может быть, это зависит от энергичной деятельности их фагоцитов, которые разрушают микробов, как только они войдут внутрь губки, а может быть, вследствие недостаточности наших знаний по этому вопросу.
Губки представляют аналогию с простейшими и миксомицетами в том, что во всех этих организмах пищеварительные и выделительные функции играют роль в реакции против посторонних тел, вредных для организма.
В губке или в пласмодии эта реакция состоит только в захватывании попавшего тела и в переваривании его, если это возможно, или, в обратном случае, в его выбрасывании. У миксомицетов эта функция принадлежит всей протоплазме, тогда как у губок она сконцентрирована в мезодерме и отчасти в эктодерме. Наружная оболочка, или вообще эктодерма, служит организму защитой против всякого рода вредных для него факторов. Плоские клетки, составляющие эктодерму, сократимы и обладают чувствительностью. Они сокращаются и открывают поры, чтобы впустить окружающую жидкость внутрь губки, если только вода не содержит ничего вредного для ее организма.
Уж давно было замечено, что для того, чтобы лучше исследовать проникновение окрашенных зерен внутрь клеток губки, надо наблюдать голодных индивидуумов. Но как только губка уже достаточно насыщена маленькими зернами, принесенными водой, поры ее более не открываются и, так сказать, не дают доступа излишку этих тел.
По наблюдениям Ленденфельда, губки держат закрытыми свои поры и мешают, таким образом, вхождению вредных тел не только в том случае, когда эти тела находятся в виде зерен, плавающих в воде, но и тогда, когда они в ней растворены. Из всех употребленных им тел (крахмал, кармин, молоко) только молоко проникало беспрепятственно с самого начала внутрь губки; для кармина поры сначала оставались закрытыми, но потом скоро открывались. Растворы многих ядовитых веществ, например морфина, вератрина или стрихнина, заставляли сжиматься поры и проникали внутрь губки только по прошествии некоторого времени.
297
В этих явлениях, обнаруживаемых эктодермическими клетками, которые сократимы, но не представляют из себя фагоцитов, можно установить аналогию и, с другой стороны, различие с пласмодиями миксомицетов. Аналогия состоит в чувствительности к химическому составу окружающей среды, что свойственно как эктодермическим клеткам губок, так и пласмодию. Разница сказывается в способе реагирования.
Пласмодий — подвижная колония клеток — удаляется от причины, вызывающей в нем отрицательную чувствительность (химиотаксию, термотаксию и т. п.); губка же — организм неподвижный — избегает той же самой вредной причины, не допуская ее проникнуть в свое тело.
Несмотря на недостаточность наших сведений, мы все-таки имеем право утверждать, что клетки губок играют активную роль в борьбе против различных вредных влияний. Особенно же важны свойства чувствительности и сократимости эктодермы, с одной стороны, а с другой — способность клеток мезодермы и энтодермы захватывать и переваривать посторонние тела. Эти факты могут служить точкой отправления для более сложных явлений реакции, существующих у других животных.
ЛЕКЦИЯ ПЯТАЯ
Coelenterata, иглокожие и черви. — Повреждения и восстановление гидр. — Скопление фагоцитов у медуз акалеф. — Фагоциты морских звезд. — Воспаление у Bipinnaria. — Реакция со стороны перивисцеральных клеток кольчатых червей. — Фагоцитная реакция в инфекциях у Nais и дождевых червей. — Борьба между фагоцитами дождевого червя и Rhabditis. Микробные инфекции червей
Coelenterata отличаются от губок более высокой организацией, но, несмотря на это, среди них есть многочисленные представители, состоящие только из двух пластов; мезодерма у них совершенно отсутствует. Мы видели на губках, что мезодерма играет главную роль в патологических процессах; интересно знать, — как происходят эти явления у животных только с двумя зародышевыми пластами, как, например, у гидры и ее родичей.
Уже в прошлом столетии часто наблюдали за тем, что делается с пресноводным полипом вследствие различного рода повреждений. Особенно благодаря Трамбле убедились в поразительной способности этого животного к полной регенерации. Можно разрезать гидру на несколько кусков, ввести в нее занозы и вообще вызвать самые серьезные повреждения, и все это не помешает гидре быстро и вполне восстановить целость своего организма. По наблюдению Ишикавы *, передняя часть поврежденной гидры совершенно восстанавливается через 20 минут. Гидры, разрезанные вдоль всего тела и растянутые на пробке, способны оправляться немного более чем через сутки. Ишикава в одном своем опыте отрезал сначала голову гидре со всеми ее щупальцами, затем разрезал в длину все ее тело, после чего поместил животное на кусок губки, вывернув энтодерму наружу (рис. 28, 1). Для повреждения энтодермы гидра, приготовленная таким образом, была вынута из воды и оставлена на воздухе в продолжение 5 минут. После этого ее сняли с пробки и положили снова в воду; гидра свернулась сначала в цилиндр, наружная сторона которого состояла из энтодермы; скоро, однако, она перевернулась таким образом, что оба пласта клеток заняли свое нормальное положение. Но во время перевертывания в гидру попала нить водоросли и помешала слиянию краев (рис. 28, 2). Тогда гидра изменила свое положение и в конце концов превратилась в закрытый мешок (рис. 28, 3), на котором появился рот со щупальцами
* Zeitschr. f. wiss. Zool., т. XLIX, 1889, стр. 433.
299
(рис, 28, 4 и 5), и она снова стала вполне нормальной гидрой спустя 6 дней после начала опыта.
Уколы и другие повреждения, сделанные инструментами, заживлялись необыкновенно быстро без скопления фагоцитов в поврежденном месте. Хотя в данном случае и не было скопления фагоцитов вследствие отсутствия мезодермы, но не надо, однако, думать, чтобы явление фагоцитоза отсутствовало здесь совершенно. Вся энтодерма гидры состоит из неподвижных фагоцитов в виде эпителиальных клеток. Они способны выпускать амебовидные отростки на свободной поверхности и захватывать ими различные посторонние тела.
У колониальных морских гидрополипов не только энтодерма, но иногда также и эктодерма состоит из фагоцитов, выполняющих важную профилактическую роль *. Эти животные способны, подобно гидре, регенерироваться. Если отрезать голову гидроиду, например, у Podocoryne, и оставить туловище в сообщении со всей колонией, то на нем появляется новая голова, а отрезанная приобретает новое туловище.
Общее во всех этих явлениях состоит в необыкновенно быстром и широком возрождении поврежденных мест, так что опасность инфекции становится ничтожной. Мы видим здесь только восстановительную сторону воспалительного процесса, но не самое явление воспаления или, по крайней мере, не скопление фагоцитов в поврежденном месте. Это последнее явление совсем, однако, не чуждо организму Coelenterata. Большинство этих животных, как, например, Medusae acraspedae, Ctenophorae и настоящие полипы, имеют достаточно развитую мезодерму, в которой встречаются амебовидные клетки, обладающие всеми свойствами фагоцитов.
Возьмем большую медузу, известную под именем Rhizostomum Cuvieri, и введем в ее студнеобразный колокол занозу, например, кусок дерева или просто булавку. На другой же день мы увидим простым глазом белое пятно вокруг постороннего тела. Наблюдение под микроскопом показывает, что это есть множество амебовидных клеток, собравшихся в месте

Рис. 28. Регенерация гидры (по Ишикава).
* См. мою работу в Arbeiten d. zool. Inst. d. Univ. zu Wien, т. V, 1883, стр. 143—146 163
300
повреждения. То же самое явление происходит у другой акалефы, Aurelia aurita. Если тело, введенное в колокол медузы, было опущено сначала в красящий порошок (например, кармин), то фагоциты, собранные на месте повреждения, оказываются наполненными красящими зернами. Амебовидные клетки, скопившись вокруг постороннего тела, остаются изолированными друг от друга или сливаются для образования маленьких пласмодиев.
Итак, мы видим, что у медуз, совершенно лишенных кровеносных сосудов, мезодермические фагоциты приближаются к постороннему телу через студенистую массу, иногда довольно твердую (как у Rhizostomum). Они захватывают введенные тела или, если эти последние очень велики, то только обволакивают их.
Аналогия этих явлений с реакцией у губок неоспорима, а между тем есть значительная разница в мезодерме губок и медуз. У губок мезодермические фагоциты играют важную роль в пищеварении твердых тел; у медуз же и у всех Coelenterata, несмотря на присутствие мезодермы, пищеварение совершается исключительно энтодермой. Энтодерма у Coelenterata состоит из фагоцитного эпителия, совершенно отделенного от мезодермы, по крайней мере у взрослых форм. Однако, хотя мезодермические фагоциты и не имеют пищеварительной функции, они все-таки сохраняют способность приближаться к посторонним телам, захватывать их или окружать и даже переваривать некоторые из них. Они проявляют эту способность не только по отношению к посторонним телам, появляющимся в теле Coelenterata при повреждениях, но также и по отношению к элементам самого животного. Так, например, неразвившиеся половые клетки, часто наблюдаемые у медуз, содержимых в неволе, постоянно становятся добычей фагоцитов, образующих вокруг них род фолликулы. Очевидно, что эти мезодермические клетки вовсе не потеряли своей первичной способности внутриклеточного пищеварения, и, хотя они вполне отделены от энтодермы, тем не менее их общее происхождение может быть доказано эмбриологическими фактами.
Развитие амебовидных мезодермических клеток на счет энтодермы — факт, очень распространенный в животном царстве. Он может быть легко наблюдаем у различных представителей иглокожих, особенно у ежей и морских звезд. Возьмем, например, Astropecten pentacantus — морскую звезду, часто встречаемую в Триестском заливе. Разделившееся яйцо превращается в шар, состоящий из мерцательных клеток. Часть их углубляется в полость зародыша для образования первого зачатка пищеварительного канала и его придатков. Личинка принимает скоро характер настоящей гаструлы и состоит уже из эктодермы — наружного слоя и энтодермы — внутреннего; она имеет форму мешка с отверстием на нижнем конце. Пространство между
301
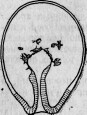
Рис. 29. Образование фагоцитов у личинки Astropecten.
двумя слоями, или общая полость личинки наполнена однородным, почти жидким веществом, которое населяется амебовидными клетками мезодермы.
Эти последние не что иное, как клетки энтодермы, отпавшие и сделавшиеся блуждающими * (рис. 29). Едва отделившись и попав в общую полость, эти подвижные клетки уже выполняют свою фагоцитную функцию. Между многочисленными личинками Astropecten, плавающими по поверхности моря, встречаются такие, у которых тонкая эктодерма поранена колючим телом, которое и могло таким образом проникнуть в общую полость тела (рис. 30). Тотчас же вслед за повреждением мезодермические клетки приближаются к постороннему телу и вполне его окружают, сливаясь в маленькие пласмодии (рис. 31). Эти последние заключают внутри себя некоторое количество ядер, вид которых доказывает полное отсутствие клеточного размножения. (Эти ядра очень заметны при действии 1/2% осмиевой кислоты и при окрашивании пикрокармином). Реакция этих личинок, которую можно проследить шаг за шагом вследствие их прозрачности, состоит исключительно в скоплении мезодермических фагоцитов вокруг постороннего тела. Надо заметить, что эти личинки совершенно лишены нервной, сосудистой и мускульной систем, так что участие этих последних в реакции должно быть исключено. Реакция, следовательно, совершается фагоцитами вполне самостоятельно и не сопровождается ни притоком жидких частей (которые совершенно отсутствуют), ни явлением размножения клеток. Отсутствие последнего легко объясняется тем, что постороннее тело, будучи очень тонким, вызвало только самое незначительное повреждение эктодермы. У более развитых и сложных личинок реакция совершается таким же образом. Я часто видел, как морская водоросль из рода Chaetoceros, снабженная очень тонкими волосами, прикреплялась к поверхности Bipinnariae (личинка Astropecten) и проникала внутрь их тела. Во всех случаях
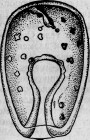
Рис. 30. Гаструла с посторонним телом в общей полости.
* См. мою работу в Zeitschr. f. wiss. Zool., т. XLII, 1885. Смотри по этому вопросу спор между г. Зеленкой и мной, а также более недавнюю работу г. Коршельта в Zoologische Jahrbucher, т. IV, 1889.
302
повреждение сопровождалось скоплением мезодермических фагоцитов и образованием пласмодиев.
В вышеприведенных случаях личинки слишком малы, чтобы служить для опыта, и приходится довольствоваться наблюдением повреждений, происшедших естественным путем, без вмешатель-

Рис. 31. Постороннее тело, изображенное на рис. 30, окруженное пласмодием личинки (сильное увеличение) .
ства человека. Но если обратиться к большим личинкам, описанным под именем Bipinnaria asterigera и представляющим также стадию развития морской звезды, то на них можно легко изучить явления реакции, вызванной искусственными повреждениями *. Надо только ввести под кожу такой личинки маленькую стеклянную трубочку или шип розы, чтобы увидеть, как мезодермические амебовидные клетки сгруппировываются вокруг введенного тела. Они образуют большие скопления, заметные для простого глаза. Все маленькие зерна, прилипшие к введенному телу, например зерна индиго или кармина, если предмет был в них помещен, жадно поглощаются фагоцитами мезодермы (рис. 32).
Если вместо твердых тел ввести под кожу Bipinnaria каплю крови, то и она окружается мезодермическими
клетками, образующими вокруг нее настоящие пласмодии, т. е. протоплазматические массы со многими ядрами, происшедшие от слияния фагоцитов (рис. 33). Изменения красных кровяных шариков внутри мезодермических клеток личинки совершенно

Рис. 32. Скопление фагоцитов вокруг занозы Bipinnaria asterigera.
* См. мою работу в Arbeiten d. zool. Inst. d. Univ. zu Wien, т. V, 1883, стр. 141 164.
303
соответствуют внутриклеточному пищеварению, которое одинаково наблюдается и по отношению жировых капель молока.
Бактерии, введенные под кожу Bipinnaria, также захватываются клетками мезодермы. Благодаря прозрачности личинок иглокожих можно легко видеть, как амебовидные клетки поглощают живых бактерий, так как последние еще подвижны.
При сравнении явлений реакции у губок, у Coelenterata, имеющих мезодерму, и у иглокожих поражаешься их сходству в существенных чертах, несмотря на различие этих трех типов. У губок мы находим мезодерму, богатую подвижными клетками, которые принимают большое участие в пищеварительной функции этих животных. Пища, попавшая в их тело, всегда проникает
в мезодерму, находящуюся в близком соприкосновении с эктодермой. У акалеф и других Coelenterata с тремя пластами мезодерма сообщается непосредственно с энтодермой только в период развития. По окончании этого периода мезодерма окончательно отделяется от энтодермы и не принимает больше участия в пищеварении, которое совершается исключительно фагоцитами энтодермы 165. У личинок иглокожих происходит также окончательное разделение двух пластов: мезодерма и здесь не участвует в пищеварении животного; энтодерма — единственный пищеварительный орган — тоже неспособна к внутриклеточному пищеварению. Последнее совершается с помощью неорганизованных ферментов, выделяемых энтодермическими клетками во внутреннюю полость.
Но, несмотря на все это различие организации, клетки мезодермы сохраняют свою способность приближаться к посторонним телам, захватывать их и переваривать, если они переваримы. Различные повреждения, произведенные этими телами, вызывают у всех описанных животных скопление мезодермических фагоцитов. Последние часто образуют при этом пласмодии, или гигантские клетки.
Общее у всех этих животных выражается в том, что их мезодермические фагоциты появляются в виде звездчатых клеток соединительной ткани; эти клетки разбросаны в полужидком или студенистом межклеточном веществе. Во всех рассмотренных случаях не было ни крови, ни плазмы, ни кровяных шариков, ни
304
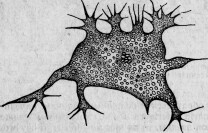
Рис. 33. Пласмодий, образованный фагоцитами бипиннарии.
кровеносных сосудов. Этих органов нет ни у губок, ни у Coelenterata, и появляются они только у иглокожих, в период гораздо более поздний, чем те, которые послужили в приведенных выше патологических исследованиях.
Если обратимся к типу столь разнообразных червей, то мы увидим сначала явления реакции, вполне примыкающие к тем, которые мы уже видели у других животных. Возьмем Mesostomum Ehrenbergi (из Turbellaria) как представителя низших червей и произведем повреждение в какой-нибудь части его тела. По прошествии некоторого времени мезодермические фагоциты соединяются в поврежденном месте. Мы увидим, что они наполнены зернами, и это делает их очень похожими на клетки кишечного эпителя, представляющего у Turbellaria настоящих фагоцитов. Мезодермические клетки представляют собой подвижные амебовидные элементы, разбросанные в межклеточной студенистой массе, составляющей соединительную ткань. Эта последняя напоминает мезодерму губок, медуз и иглокожих.
У более высших червей мезодермические фагоциты являются в виде клеток, плавающих в перивисцеральной жидкости, или в виде перитонеального эндотелия. Эти два рода клеток имеют одинаковые фагоцитные свойства, очень ясно выраженные. Аналогия в их функциях может объяснить тот факт, что у очень близких видов перивисцеральные клетки то являются очень развитыми, то вовсе отсутствуют. Эти мезодермические элементы служат, кроме того, органами дыхания и выделения *.
Заноза, введенная в перивисцеральную полость кольчатого червя, например Terebella, скоро покрывается толстым слоем «лимфатических» клеток. Фагоцитные свойства этих последних доказываются той легкостью, с которой они захватывают мелкие зерна (окрашенный порошок или что-нибудь другое), прилипшие к занозе. Интерес этого факта увеличивается присутствием у большинства кольчатых червей очень развитой и вполне замкнутой сосудистой системы. Реакция против посторонних тел совершается у них исключительно мезодермическими фагоцитами; кровеносные же сосуды остаются совершенно пассивными. Это можно доказать, благодаря довольно сильной окраске кровяной жидкости.
Те же самые явления наблюдаются у кольчатых червей с развитой сосудистой системой, но не имеющих перивисцеральных фагоцитов. При исследовании Nais proboscidea попадаются иногда особи, зараженные личинками Gordius. Эти личинки, проникнув в общую полость тела, возбуждают фагоцитную реакцию, производимую единственно перитонеальными клетками. Они посылают протоплазматические отростки, образуя род пласмодия вокруг личинок. Эти последние защищаются, выделяя хитинную
* См. Grobben. Die Pericardialdruse der chaetopoden Anneliden. Sitzungsber. d. K. Akad. d. Wiss. zu Wien, т. LCVII, 1888.
305
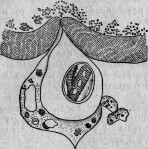
Рис. 34. Личинка Gordius, закистированная и окруженная пласмодием Nais.
оболочку и покрываясь кистой. Иногда эти кисты лежат очень близко от сосудов, которые не оказывают по отношению к ним никакой реакции (рис. 34). Если бы содержимое сосудов проникало в полость пласмодической оболочки, то это легко можно было бы заметить, так как кровяная плазма окрашена в желтый цвет, а перивисцеральная жидкость совершенно бесцветна.
Чтобы убедиться в фагоцитных свойствах перитонеальных клеток Nais proboscidea, надо наблюдать индивидуумов, зараженных микроспоридиями, принадлежащими к микробам Pebrina 166. (Поранить искусственно этих кольчатых червей невозможно вследствие их очень малых размеров.) Эти паразиты вызывают также реакцию со стороны перитонеального эндотелия, и маленькие споры захватываются тогда внутрь фагоцитов, отделенных от брюшины. Иногда эти споры окружаются вакуолями совершенно так, как в типичных случаях внутриклеточного пищеварения. Кольчатые черви больших размеров могут также служить для подобных исследований. Очень полезно в этом отношении изучать обыкновенных дождевых червей, организм которых часто бывает вместилищем многих паразитов. Из этих последних чаще всего встречаются грегарины из рода Monocystis; они паразитируют в мужских половых органах. Появляясь в них, эти подвижные животные встречают много амебовидных клеток, принадлежащих к категории наиболее деятельных фагоцитов. Последние снабжены тонкими или лопастными протоплазматическими отростками и легко захватывают все попадающиеся им посторонние тела (рис. 35, А). Даже на препарате, в водяной влаге кролика или другой какой-нибудь безвредной жидкости, эти клетки проявляют свою фагоцитную деятельность: они пожирают зерна краски или какие-нибудь другие маленькие тела, находящиеся в препа-
306
Рис. 35. А — фагоцит дождевого червя; В — скопление фагоцитов дождевого червя вокруг постороннего тела.
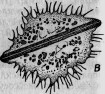

рате. Встречая предметы большей величины, например растительные волокна, фагоциты соединяются группами и все-таки окружают эти тела своей протоплазмой (рис. 35, В). Те же самые клетки реагируют и против паразитов, проникших внутрь дождевых червей. Пока грегарины находятся в деятельной стадии, они отталкивают фагоцитов резкими движениями, так что только в редких случаях эти клетки могут прикрепляться к телу паразита. Но как только грегарины переходят в покоящееся состояние, фагоциты приклеиваются к их поверхности и образуют иногда вокруг них толстую массу. Паразит, стесненный этой живой оболочкой, защищается, как и личинки Gordius, выделяя кисту.
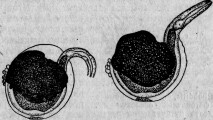
Рис. 36. Живой Rhabditis, окруженный массой фагоцитов дождевого червя.
Защищенная таким образом грегарина начинает производить споры, разделяясь на большое число все более и более мелких сегментов и образуя, наконец, pseudonavicellae167. Фагоциты между тем продолжают оказывать свое влияние на паразита и часто даже убивают его. Закистированная грегарина продолжает защищаться, выделяя хитин. Это вещество получается в очень большом количестве и придает паразиту чрезвычайно неправильное и оригинальное очертание.
Случается, что содержимое грегарины, окруженной фагоцитами, становится сильно преломляющим свет, и паразит умирает, побежденный своим врагом.
Сами фагоциты, окруженные паразитами, тоже претерпевают значительные изменения; они превращаются в плоские клетки, спаянные между собой и неподвижные.
Иногда такая капсула, представляющая строение соединительной ткани, остается очень тонкой, но чаще всего она утолщается вследствие появления новых слоев клеток. В числе этих последних встречаются элементы, очень богатые бурым пигментом.
Во время борьбы паразита с фагоцитами червя кровеносные
307

Рис. 37. Rhabditis, освобожденный от своей фагоцитной оболочки с целью показать кутикулярные придатки.
сосуды этого последнего остаются вполне бездеятельными, несмотря на свое очень значительное развитие. Они не представляют ни видимого изменения в объеме, ни выделения красноватой плазмы.
Нематоды из рода Rhabditis довольно часто проникают в семенные железы и в общую полость тела дождевого червя. Эти животные, несмотря на всю свою подвижность, крепость кутикулы и довольно большую величину, должны тем не менее вести борьбу с многочисленными фагоцитами земляного червя. Фагоциты окружают свернувшуюся нематоду плотной массой, подобной оболочке, образованной вокруг грегарины. Микроскопическое наблюдение доказывает прежде всего, что фагоциты окружили живого врага, так как он еще движется внутри фагоцитной массы
(рис. 36). Этот червь защищается выделением хитиновых слоев, никогда не образующих настоящей кисты, а только добавочную оболочку, часто необыкновенно толстую. Подобное обильное выделение истощает паразита, так как он при этом постепенно лишается жировых капель, которыми вначале был наполнен, и делается вполне прозрачным.
Хитиновые слои становятся все толще и толще и в конце концов образуют неправильные придатки, придающие Rhabditis странную и неправильную форму (рис. 37). Отделяя фагоцитные массы от червя, окруженного добавочной кутикулой, часто можно видеть, что эта немотода высвобождается, оставляя свою оболочку внутри слоя фагоцитов.
С другой стороны, при исследовании содержимого мужских половых органов дождевого червя находят иногда внутри фагоцитных капсул блестящие массы, в которых легко узнать совершенно обесформленные кутикулярные оболочки и остатки нематоды, замурованной в продукты своего выделения.
Мы видели здесь настоящую борьбу двух живых существ, принадлежащих к одному типу животного царства. Нематода защищается с помощью кожных выделений, земляной же червь борется целой армией подвижных клеток, обладающих фагоцитными свойствами.
Очевидно, что фагоциты, окружая паразита, мешают ему жить, хотя мы еще не можем точно определить самой сути этой фагоцитной реакции. Может быть, фагоциты мешают притоку питательных веществ или кислорода, может быть, они выделяют какое-нибудь вредное вещество. Эти сложные вопросы могут быть выяснены только тщательными исследованиями,
308
к которым можно будет приступить, имея уже пригодные для этого методы.
В настоящее же время мы должны ограничиться констатированием факта, что земляные черви, как и кольчатые вообще, реагируют против различных инфекций с помощью фагоцитов перивисцеральной жидкости, а не посредством крови или хорошо развитых кровеносных сосудов.
Эта реакция совершается одинаково как против грегарин, так и против нематод.
Относительно последних надо заметить важный факт отсутствия в их организме блуждающих клеток. Фагоцитная система нематод сводится, вероятно, к очень оригинально развитым мускульным фагоцитам. Животные эти защищаются своими кутикулярными выделениями, представляющими значительное сопротивление. В этом отношении нематоды напоминают растения, клетки которых защищаются толстыми и твердыми оболочками, и, так же как и растения, они чаще всего подвергаются нападению паразитических грибов. Эти последние обладают, во-первых, громадной силой роста и, кроме того, выделяют ферменты, способные растворять вещество самых твердых оболочек, как, например, клетчатку. Между заразными болезнями нематод очень распространена одна, вызываемая паразитизмом одного из мукоров (Mucor helminthophorus de Вагу), наводняющего кишечный канал и половые органы кошачьей аскариды (Ascaris mystax)*. Кроме того, у нематод есть много других заразных болезней, вызываемых различными представителями класса грибов. Наиболее замечательный есть, без сомнения, Arthrobothrys oligospora Fres.; по наблюдениям Цопфа**, эта плесень ловит Anguillula посредством петель и затем выпускает нити в тело нематод.
Проникши сюда, гриб свободно развивается в общей полости и причиняет смерть животному, сопровождающуюся полным жировым перерождением. В конце концов от нематод остается только кутикула и хитиновая оболочка мужских половых органов.
Кроме эпидемий, производимых настоящими грибами, нематоды заражаются еще хитридиями, а также низшими организмами, близкими к тем, которые вызывают болезнь ядра и ядрышка у Paramaecium***.
Обзор патологических явлений у Coelenterata, иглокожих и червей показал нам, что некоторые из этих животных реагируют главным образом очень быстрой регенерацией поврежденных частей; другие же защищаются выделением хитиновых слоев. Эти два способа защиты представляют только частные случаи, тогда
309
* Zeitschr, f. wiss. Zool., т. II, 1862, стр. 135.
** Nova Acta Acad. Leopold, т. XLVII, стр. 167, и Pilze, 1890, стр. 240. *** См. Бютчли. Studien uber die ersten Entwickelungsvorgange der Eizelle, Frankfurt, 1876, стр. 360.
как борьба с помощью амебовидных и подвижных клеток, окружающих или захватывающих постороннее тело, составляет общее правило с редкими исключениями. Реакция фагоцитных клеток совершается в силу их чувствительности, без вмешательства нервной или сосудистой систем. Во всех вышеприведенных случаях фагоциты являлись или в виде подвижных клеток соединительной ткани, или в виде клеток перивисцеральной полости. До сих пор мы еще не видели примеров фагоцитной деятельности кровяных шариков. Правда, в приведенных случаях мы имели дело только с животными, лишенными форменных элементов крови. Кольчатые черви, у которых уже есть белые кровяные шарики, плавающие в сосудах, встречаются редко, и к тому же их кровяные лейкоциты немногочисленны, и их гораздо меньше, чем в перивисцеральной полости.
ЛЕКЦИЯ ШЕСТАЯ
Суставчатоногие, мягкотелые и оболочниковые. — Их кровеносная система. — Фагоциты этих беспозвоночных. — Селезенка моллюсков. — Воспалительная реакция. — Нормальный диапедез у асцидий. — Введение бактерий в тело асцидий и ракообразных. — Инфекционная болезнь у Talitrus. — Болезни дафний. — Внедрение бактерий в тело насекомых. — Эпидемии у насекомых
Многие беспозвоночные имеют кровяные тельца в форме бесцветных клеток, циркулирующих в жидкости. Эта жидкость приводится в движение сердцем, которым они всегда снабжены. У всех суставчатоногих, моллюсков, а также у оболочниковых общая полость тела соединена с сосудистой полостью. У низших представителей этих типов мы находим единственный орган кровеносной системы — сердце. (Мы оставляем в стороне некоторые группы, не имеющие даже следов сосудистой системы, как многие Copepoda, Ostracoda и др.). Это сердце имеет форму мешка или трубки, открытой на концах для выхода крови и снабженной боковыми отверстиями, через которые жидкость проникает в сердце. К этому центральному органу присоединяется прежде всего одна или несколько главных артерий, открывающихся в систему полостей, где циркулирует кровь до входа в сердце. У более развитых беспозвоночных, особенно у мягкотелых, присоединяется еще венозная система, иногда очень развитая, как, например, у головоногих. Но всегда, без исключения, даже в случае образования многочисленных сосудистых разветвлений, между артериальной и венозной системой находится сеть лакун — остаток общей полости, — наполненных кровью.
Кровяные тельца представляют собой бесцветные клетки (за редкими исключениями) с одним, редко двумя ядрами и с протоплазмой, способной к амебовидным движениям. У многих беспозвоночных эти кровяные тела состоят из одного только рода подвижных клеток с незначительной зернистостью. У других же, как, например, у многих насекомых и моллюсков, есть два рода лейкоцитов: зернистые с большим количеством крупных зерен и гиалиновые с немногими зернами или совсем без них. Нас особенно интересует последняя категория лейкоцитов.
Лейкоциты суставчатоногих, мягкотелых и оболочниковых представляются вообще амебовидными фагоцитами: они отличаются от белых кровяных шариков позвоночных присутствием одного круглого или овального, не лопастного ядра. У интересующих нас беспозвоночных совсем нет многоядерных лейкоцитов, точно так же, как нет у них вполне замкнутой сосудистой
311
капиллярной системы. Лейкоциты трех вышеназванных типов беспозвоночных обнаруживают ясные фагоцитарные свойства. В 1862 г. именно у одного из представителей этих беспозвоночных было сделано в первый раз открытие свойства лейкоцитов захватывать посторонние тела. Впрыскивая индиго в моллюска Tethys, Геккель * наблюдал, что зерна этой краски находились внутри кровяных телец. Опыты с некоторыми другими родами животных доказали, что этот факт имеет общее значение, что и было подтверждено впоследствии несколькими наблюдателями. Вследствие этого удивительно, что Грисбах ** высказал недавно сомнение по поводу фагоцитоза белых кровяных шариков безголовых моллюсков 168. Грисбаху не удалось констатировать захватывания лейкоцитами впрыснутого порошка, и поэтому он думает, что при нормальных условиях фагоцитоза у безголовых моллюсков не происходит. Весьма вероятно, что неудовлетворительные результаты у этого автора получились вследствие слишком сильного разбавления порошка водой, что должно было вызвать разбухание лейкоцитов. Производя более тщательно опыты, можно легко доказать, что лейкоциты моллюсков, как и многих других животных, жадно захватывают твердые тела, с которыми они приходят в соприкосновение. Особенно удобно производить эти опыты с прозрачными моллюсками, как Phylliroё, которую в живом состоянии можно наблюдать под микроскопом.
У некоторых гастеропод существует, кроме белых кровяных шариков, еще одна особая форма фагоцитов, составляющих у этих беспозвоночных род селезенки. Это важное открытие было недавно сделано Ковалевским***. Он доказал, что твердые тела, впрыснутые в кровь Pleurobranchia и некоторых других родов (Philine, Gasteropteron, Doris), скопляются в особом органе, открытом Лаказом-Дютье и описанном им под именем «неопределенной железы». Клетки этой селезенки захватывают и переваривают большое количество посторонних тел (кровяные шарики, желточные зерна, молочные тельца), как это было вполне установлено Ковалевским.
Фагоциты, столь распространенные у вышеназванных беспозвоночных, реагируют против всяких повреждений, искусственных или случайных. Очень часто можно встретить прозрачных ракообразных, например дафний или бранхипуса, у которых стенки тела имеют бурые пятна, происходящие от укушения других особей. Под этими царапинами находят обыкновенно скопление лейкоцитов, остающихся в поврежденном месте до полного заживления раны****. Производя у подобных животных осторожно слабое повреждение, можно видеть под микроскопом, как лейко-
* Die Radiolarien, 1862, стр. 104. ** Archiv f. mikrosk. Anat., т. XXXVII, стр. 86. *** Зап. Новоросс. О-ва естествоисп., т. XV, 1890.
**** Очень быстрое возрождение эпидермиса у суставчатоногих способствует быстрому заживлению их ран.
312
Рис. 38. Хвостовой придаток Argulus в состоянии воспаления.

циты направляются к поврежденному месту, где и фиксируются.
Очень удобным объектом для такого рода опытов могут служить хвостовые придатки Argulus; искусственное повреждение вызывает у них быстрое скопление лейкоцитов (рис. 38).
Довольно объемистые посторонние тела (например, всякого рода занозы) могут быть введены в личинок разных жуков (майский жук, Oryctes и др.), моллюсков, как, например, Thethys или Phylliroё, и асцидий*. Во всех этих случаях большое количество лейкоцитов собирается вокруг постороннего тела, захватывая введенные маленькие зерна (например, кармин, которым была
натерта заноза).
Итак, постоянно происходит фагоцитная реакция, проявляющаяся то в форме инфильтрации лейкоцитов вокруг постороннего тела, то в образовании капсулы,
состоящей из массы лейкоцитов. Эта экссудативная и воспалительная реакция, часто сопровождающаяся образованием гигантских клеток, не имеет никакого отношения к диапедезу по той простой причине, что членистоногие и моллюски имеют не вполне замкнутую сосудистую систему, но соединенную с общей полостью тела.
Есть только один пример диапедеза у беспозвоночных, и этот случай так интересен, что заслуживает особого внимания. Асцидии покрыты очень толстой оболочкой, расположенной вне эпидермиса. Эта оболочка состоит из целлюлозного вещества и заключает большое количество амебовидных клеток, снабженных подвижными придатками. Так как этот слой со своими клетками находится вне эпидермиса, то на него смотрели, как на кожное выделение, заключающее клетки эктодермического происхождения. Последние исследования Ковалевского** доказали, что эта теория не имеет никакого основания и что в действительности клетки оболочки асцидий не что иное, как лейкоциты, прошедшие сквозь эпидермис животного. Эти клетки мезодермического происхождения — очень деятельные фагоциты, способные захватывать всякого рода твердые тела и в том числе атрофирующиеся органы. Введение заноз в оболочку асцидий вызывает скопление этих фагоцитов, производящих, таким образом, род инфильтрации оболочки.
Мы видели здесь пример того, что диапедез через эпидермис совершается вполне нормальным образом и совершенно независимо от воспаления. Это последнее происходит с помощью тех же фагоцитов, собирающихся вокруг посторонних тел.
* См. Arbeiten. d. zool. Inst. d. Univ. zu Wien, т. V, 1883, стр. 153. ** Отчет О-ва петерб. естествоисп., ноябрь, 1890.
313
Любарш* подтвердил, что подвижные клетки оболочки асцидий собираются массами вокруг посторонних тел, введенных через укол. Он был менее счастлив в своих опытах с сибиреязвенными бактериями, привитыми в организм различных асцидий.
Только часть бактерий, введенных в оболочку, была захвачена; другие же, оставаясь вне фагоцитов, представляли явные признаки перерождения. Любарш не исследовал прямого влияния жидких частей оболочки на бактерий, вследствие чего его опыты, к тому же немногочисленные, не позволяют сделать никакого решительного заключения. Объясняя их себе, не надо забывать, что они были произведены в марте, когда низкая температура могла иметь вредное влияние на лейкоцитов. Кроме того, оболочка асцидий должна вообще представлять мало благоприятную среду для развития бактерий и для произведения ими токсинов**.
Любарш*** сделал также несколько аналогичных опытов с «морскими ракообразными» и не получил лучших результатов. Эти опыты были описаны так поверхностно, что критика их невозможна. А между тем существует много фактов, доказывающих самым очевидным образом фагоцитные свойства лейкоцитов различных ракообразных.
Герману и Каню**** удалось ввести паразитного гриба (близкого к Oidium) в полость тела Talitrus и производить почти всегда смертельную болезнь этих ракообразных. Развитие паразита вызывает со стороны организма реакцию, которая проявляется ясно выраженным фагоцитозом лейкоцитов. «Ha 7-й день, — говорят вышеназванные авторы, — кровь, до тех пор прозрачная, становится мутной. Муть увеличивается в продолжение 8-го и 9-го дня по мере размножения паразитов. В продолжение этого периода замечается особенно деятельный фагоцитоз: при рассматривании с сильным увеличением крови, фиксированной осмиевой кислотой и покрашенной пикрокармином, видны микроорганизмы, заключенные в различном количестве (от 1—20 и более) в кровяных шариках. Они претерпевают внутри протоплазмы последовательные фазы внутриклеточного пищеварения; они становятся там более бледными, теряют блеск и увеличиваются в объеме, вследствие, главным образом, разбухания их оболочки. Наконец место, занимаемое ими, обозначается только бесцветной вакуолей, сохраняющей в продолжение некоторого
* Untersuchungen uber die Ursachen der angeborenen und erworbenen Immunitat, Berlin, 1891, стр. 75.
** Я должен здесь напомнить, что у сложных асцидий, как Bothryllus, фагоциты оболочки очень часто бывают наполнены различными бактериями. Это бывает у только что пойманных в море животных.
*** Untersuchungen uber die Ursachen der angeborenen und erworbenen Immunitat, стр. 77.
**** Comptes rendus de la Societe de Biologie, 1891, стр. 646.
314
времени продолговатую форму паразита». Фагоцитные функции, помимо кровяных шариков, исполняются также периартериальными клетками, которые, однакоже, не в состоянии переваривать грибов. Последние, таким образом, побеждают и убивают Talitrus.
На представителях того же рода амфиподов паразитируют светящиеся бактерии, что было открыто Жиаром и Биллье*.
Многие из ракообразных подвержены различным заразным болезням, изучение которых представляет большой интерес с патологической точки зрения вообще и ввиду разрешения задачи воспаления в частности. Особенно удобны для подобного изучения дафнии по прозрачности и по малым размерам их тела, а также вследствие их частых и разнообразных болезней. У них встречаются болезни, производимые бактериями, спорозоями и сапролегниями. Но самая интересная инфекционная болезнь производится одним грибом, размножающимся почкованием (род дрожжей, описанных под именем Monospora bicuspidata**). Этот гриб нападает на Daphnia magna и находится в изобилии в Париже (бассейн рептилий в Jardin des plantes) и его окрестностях.
Среди многочисленных индивидуумов этих ракообразных встречаются экземпляры с беловатым молочным оттенком. Под микроскопом их общая полость представляется наполненной почти сплошь маленькими телами в форме тонких иголок (рис. 39), прикрепленных к стенкам сердца или плавающих в полости. Более внимательное изучение тотчас показывает, что это очень тонкие споры, лежащие внутри оболочки (рис. 40, 5).
Рядом с вполне образовавшимися спорами заметны удлиненные клетки и овальные конидии, размножающиеся, подобно дрожжам, почкованием (рис. 40, 1—4).
Дафния, наводненная массой этих грибов, всегда в конце концов погибает, причем ее труп бывает переполнен зрелыми спорами. Другие дафнии, питаясь различного рода остатками, находящимися в глубине вод, проглатывают игловидные споры и таким образом заражаются через пищеварительные органы. Во внутренностях дафнии спора освобождается от своей оболоч-
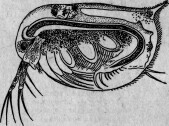
Рис. 39. Дафния, наводненная моноспорами.
* Comptes rendus de la Societe de Biologie, 1889, стр. 593. ** Virchow's Archiv, т. XCXVI, стр. 177.
315
ки, прокалывает их и проникает отчасти или вполне в общую полость тела ракообразного. Но, едва попав наружу кишечной стенки, спора подвергается нападению со стороны лейкоцитов, принесенных кровяным потоком. Эти клетки укрепляются на споре, обволакивают ее (часто сливаясь в пласмодии) и подвергают ее целому ряду замечательных превращений. Спора, захва-
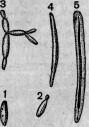
Рис. 40. Различные стадии моноспоры: 1 — молодая конидия; 2, 3 — почкующиеся конидии; 4 — удлиненная кондиция; 5 — спора.

Рис. 41. Споры моноспоры, окруженные лейкоцитами дафнии.
ченная лейкоцитами, теряет прежде всего правильность своих контуров, становится угловатой и превращается в конце концов в кучу буроватых зерен (рис. 41, 1—4), в которых нельзя было бы отличить перерожденную спору, если бы не знать их происхождения. Доказательством того, что это перерождение происходит действительно под влиянием фагоцитов, могут служить
споры, половина которых уже окружена лейкоцитами, тогда как другая половина или еще находится в кишечной стенке, или выходит наружу из кожи животного. В подобных случаях превращению подвергается только часть, окруженная фагоцитами, другая же, вне фагоцитного влияния, остается вполне неизменной (рис. 42). Когда большое количество спор проникает сразу в общую полость тела дафнии, то образуется скопление лейкоцитов, напоминающее клеточную инфильтрацию, или экссудат (рис. 43 а, b). Мы видим здесь вполне тождественное явление с тем, которое происходит вследствие описанных выше повреждений.
Фагоцитная деятельность лейкоцитов, так резко выраженная и которую так легко изучать на прозрачных дафниях, разрушает
316
Рис. 42. Удлиненная конидия моноспоры, окруженная двумя лейкоцитами.
споры патогенного микроба, мешает развитию паразита, защищая таким образом зараженный организм.
Мне случалось не раз выделять зараженных дафний и сохранять их вполне здоровыми благодаря разрушению спор фагоцитами. Если, наоборот, число проглоченных спор постоянно увеличивается или если по какой-нибудь другой причине фагоцитная защита становится недостаточной, споры начинают прорастать, образуя почкующиеся конидии. Хотя эти вегетативные стадии тоже преследуются лейкоцитами, однако дафния становится неизбежно добычей паразитов и погибает по прошествии нескольких дней. Это происходит оттого, что конидии размножаются слишком быстро и выделяют какое-то ядовитое вещество, растворяющее лейкоцитов. В конце болезни в полости тела дафнии циркулируют только конидии; фагоциты же вполне исчезли.
Совершенно неоспоримо, что вся история этой болезни дафний сводится к борьбе двух элементов: клеток паразита и фагоцитов.
Несмотря на поразительную деятельность первых, дафнии, защищаемые своими фагоцитами, одерживают в большинстве случаев победу. Вот почему в бассейне или аквариуме, где часто свирепствует болезнь monospora, количество дафний остается все-таки громадным. Несмотря на ежедневное умирание многих особей, большинство дафний хорошо противостоит этой болезни и восполняет, таким образом, число умерших.
Гораздо большие размеры принимают болезни, причиненные паразитами в случаях отсутствия фагоцитов. Примером могут служить сапролегнии. Споры этих грибов прорастают на наружной поверхности дафний или других ракообразных (например, бранхипуса) и пускают нити своего мицелия в тело этих животных.
Часто эти нити пользуются маленькими трещинами и отверстиями, произведенными различными причинами (раны или маленькие канальцы, проделанные спорами monospora). Про-
317
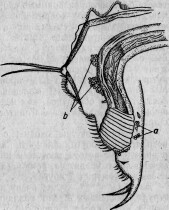
Рис. 43. Задняя часть дафнии: а, b — споры моноспоры, окруженные скоплением лейкоцитов.
никнув в полость тела ракообразных, сапролегния продолжает свое развитие в этой среде, наполненной кровью, не встречая в этом никаких препятствий. Лейкоциты проявляют полную индифферентность по отношению к развивающемуся мицелию.
Этот последний в конце концов их растворяет и ведет за собой верную смерть зараженного животного. При эпидемии сапролегний в аквариуме погибают все находящиеся в нем дафнии и бранхипусы*.
Многие другие болезни дафний, как, например, причиняемые бактерией Pasteuria ramosa** или Spirobacillus Cienkowskii***, a также болезни, производимые спорозоями (пебрина и др.), встречают очень слабое сопротивление со стороны фагоцитов, что естественно ведет к смерти животного.
Слабая фагоцитная защита, поражающая нас у ракообразных, вероятно, находится в зависимости от толщины кутикулярных стенок, одевающих не только наружную поверхность этих животных, но также и их пищеварительный канал. Эта хитинная кутикула очень тверда и не пропускает большинства микробов. Мы видим, что многие маленькие ракообразные, снабженные очень твердой оболочкой, как некоторые Copepoda, могут совершенно обходиться без фагоцитов и вовсе не имеют кровяных шариков.
Насекомые вполне сходны с ракообразными в отношении воспаления и сопротивляемости разным микробам. Всевозможные повреждения возбуждают и у них скопление фагоцитов, в чем можно легко убедиться, прижигая концы хвостовых придатков у личинок эфемерид и других.
Бальбиани**** сделал очень интересное исследование о бактериях, введенных в тело различных насекомых и пауков. Он нашел, что многие сапрофитные бактерии патогенны и даже смертельны для многих суставчатоногих. Насекомые, богатые лейкоцитами, как некоторые прямокрылые, особенно сверчки, отлично сопротивляются вхождению бацилл. Другие же отряды насекомых, отличающиеся бедностью крови и лейкоцитов (чешуйчатокрылые, двукрылые и перепончатокрылые), очень подвержены инфекции сапрофитов. «Сопротивление насекомых первой категории должно быть приписано действию на бацилл двух элементов организма: с одной стороны, кровяные шарики захватывают с помощью псевдоподий плавающих в крови бацилл; внутри кровяных шариков бациллы быстро разрушаются. С другой стороны, на бацилл действуют элементы перикардиальной ткани, состоящие из больших многоядерных клеток. Эти последние окружают сердце или спинной сосуд в виде пластинок или
* Бранхипусы и Artemia подвержены болезни, производимой monospora. Патологические явления в подобных случаях должны быть еще изучены. ** Annales de 1'Inst. Pasteur, 1888, стр. 165. *** Там же, 1889, стр. 265. **** Comptes rendus de l'Academie des Sciences, т. CIII, стр. 952.
318
более или менее развитых клеточных шнуров, смотря по типу животного. Перикардиальная ткань имеет способность удерживать бацилл, гонимых кровью, и захватывать их внутрь составляющих ее клеток, где они и разрушаются, как в кровяных тельцах» (стр. 953).
Насекомые очень редко бывают подвержены эпидемиям бактериального происхождения, хотя и чувствительны к инфекциям, производимым очень распространенными и с виду совсем безвредными бактериями. Причина заключается, вероятно, в том, что бактерии не могут проникнуть сквозь твердые кутикулярные стенки, одевающие кожу, кишечный канал и трахеи насекомых. Кроме flacherie — болезни шелковичного червя, открытой Пастером и поражающей кишечный канал 169, существуют у личинок насекомых и некоторые другие болезни, причиняемые бактериями.


Рис. 44. Лейкоцит Cleonus, взятый в двух различных фазах своего движения. Рядом — непоглощенные конидии Isaria.
Так, личинка Anisoplia austriaca, в южной России, заражается бациллой, очень напоминающей по своей удлиненной форме бациллу сибирской язвы. При начале заболевания зараженные личинки ничем не отличаются от нормальных и только после полного заражения крови они проявляют признаки болезни, за которыми скоро следует смерть.
Гораздо чаще встречаются болезни, вызываемые грибами или спорозоями, т. е. паразитами, лучше бактерий приспособленными к проникновению через хитиновые выделения насекомых. Сила прорастания грибов дает им средство проникать через кутикулярные стенки; спорозои же имеют подвижную амебовидную стадию, допускающую их в наиболее защищенные места.
Конидии грибов при размножении в крови насекомых встречают иногда некоторое сопротивление со стороны фагоцитов. Это можно заключить из наблюдения де-Бари над конидиями Cordiceps militaris, захваченного лейкоцитами гусениц. Но в большинстве случаев нити мицелия и конидии развиваются беспрепятственно в крови. Это особенно приложимо к Isaria destructor, заражающей Cleonus punctiventris в состоянии личинки, куколки или жука. Зеленая овальная спора паразита прорастает на
319
поверхности организма, выпуская маленькую нить. Эта последняя с трудом протыкает кутикулу, которая буреет вокруг укола, сделанного паразитом. Преодолевши препятствие, нить проникает в общую полость, омываемую кровью, и беспрепятственно размножается там. Лейкоциты приближаются иногда к нити или к конидиям, оторванным от этой последней, но не захватывают паразитов (рис. 44, 45). Таким образом, паразиты заражают все животное и превращают его в твердую массу, столь характерную для трупов насекомых, умерших от различного рода «мюскар-
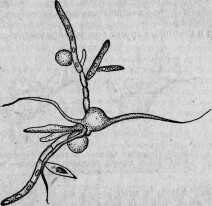
Рис. 45. Свободные конидии Isaria в самом близком соседстве с лейкоцитами Cleonus.
дин». Эти эпидемии производят часто громадные опустошения среди насекомых. Припомним потери, причиненные когда-то мюскардиной шелковичного червя. Эпидемия, производимая Isaria destructor, захватывает некоторые виды жуков, а именно Cleonus punctiventris. Часто больше половины этих насекомых, столь вредных для свекловицы, погибает от этих паразитов. Владетели свекловичных плантаций юго-западной России по смертности, причиняемой среди Cleonus «зеленой мюскардиной», делают расчет о количестве семян, нужных для посева. Они пришли к убеждению, что без естественной помощи Isaria destructor разведение свекловицы в вышеназванной местности было бы невозможно.
Болезни насекомых, причиняемые спорозоями, особенно пебрина шелковичного червя, еще не были изучены с точки зре-
320
ния сравнительной патологии воспаления. Знают*, что болезнь эта производится микроспоридией и что ее амебовидное состояние позволяет ей проникнуть во внутренность различных клеток, как, например, в молодые яйцевые клетки; но не исследовали еще вопроса о том, вступает ли паразит в борьбу с фагоцитами. У дафний, точно так же подверженных нападению микроспоридий, сопротивление лейкоцитов очень слабо и проявляется только по отношению к спорам. Амебовидная стадия развивается в самом близком соседстве с лейкоцитами, и эти последние не принимают никакого участия в ходе развития болезни. Микроспоридии, свободно развивающиеся в общей полости тела дафнии, переполняют в конце концов все животное, совсем не разрушая лейкоцитов. Последние циркулируют в крови и временно укрепляются на поверхности паразита, как на каком-нибудь совершенно безвредном теле.
Резюмируя эту главу о явлениях реакции беспозвоночных, снабженных амебовидными и фагоцитными кровяными шариками, мы должны будем заключить, что все они имеют способность производить скопление фагоцитов вокруг поврежденного места. Воспалительная реакция происходит вследствие всякого рода повреждений (прижигание, введение заноз, укусы и т. д.). Она проявляется также в различных инфекциях, как, например, у дафний, зараженных monospora. В случаях распространенного воспалительного фагоцитоза лейкоциты разносятся кровяным потоком и благодаря своей чувствительности фиксируются в месте повреждения. Полостное кровообращение облегчает приток лейкоцитов и делает бесполезным специальное приспособление для прохода этих клеток, как мы это видели у позвоночных.
Но часто лейкоциты имеют отрицательную чувствительность, что очень благоприятствует доступу разного рода паразитов. В подобных случаях главным образом хитиновые выделения, покрывающие животное, защищают его от вторжения паразитов. Мы встречаем у членистоногих средство защиты, подобное виденному нами у нематод и у растений, но только представители этого типа беспозвоночных очень редко лишены фагоцитов; в большинстве случаев членистоногие представляют уже более или менее развитую систему фагоцитной защиты.
* См. особенно Бальбиани. Lecons sur les sprozoaires, 1884, стр. 150 и cл.
ЛЕКЦИЯ СЕДЬМАЯ
Позвоночные. — Амфиоксус. — Зародыши аксолотов. — Молодые личинки хвостатых амфибий. — Сравнение с беспозвоночными. — Головастики. — Диапедез. — Блуждающие клетки. — Неподвижные клетки. Фагоцитарные свойства лейкоцитов. — Можно ли причислить неподвижные клетки к фагоцитам? — Превращение лейкоцитов в неподвижные клетки. — Участь непревращенных лейкоцитов. — Эволюция воспаления в организованном мире
Amphioxus lanceolatus, единственный из ныне живущих низших позвоночных, резко отличается в патологическом отношении от всех своих родичей. Он совершенно лишен каких бы то ни было кровяных телец и имеет только очень незначительное количество соединительнотканных подвижных клеток. Поэтому все попытки вызвать у него воспалительные явления давали отрицательный результат. Ни прижигание ляписом, ни поранение острым лезвием не вызывают видимой реакции. Это, несомненно, находится в связи с тем, что покровы амфиоксуса в высшей степени плотны и служат ему надежной защитой. В этом отношении он приближается к нематодам, к другим животным и даже растениям, выделяющим защитительные оболочки.
Чтобы встретить у позвоночных реакционные* явления, подобные тем, которые мы видели у беспозвоночных, снабженных мезодермой, надо обратиться к классу рыб. Все представители этого класса обнаруживают точно такие же воспалительные явления, как и высшие позвоночные. Но так как исследование на живых рыбах очень затруднительно, то лучше прямо перейти к амфибиям. Личинки их могут служить классическим объектом исследования. Всего лучше изучать воспалительные явления позвоночных на хвостовом плавнике личинок хвостатых (тритон и аксолот) и на головастиках бесхвостых.
Начнем наши наблюдения с хвостатых, низшей группы амфибий.
Я всегда употреблял белых аксолотов как наиболее удобных для исследования воспаления. Зачаток плавника их зародыша совершенно лишен кровеносных и лимфатических сосудов. Зачаток этот состоит, помимо эпидермиса, из слоя мезодермических клеток, которые очень часто распадаются на две категории. Одни клетки неподвижны, с разветвленными отростками в виде оленьего рога; другие же подвижны, с более толстыми отростками, мало или вовсе не разветвленными. Хотя большая часть соеди-
* Реактивные. — Ред. 322
Рис. 46. Соединительная ткань плавника зародыша аксолота: а — подвижные клетки.
нительной ткани состоит из неподвижных клеток, блуждающие клетки, однако, встречаются в довольно большом количестве (рис. 46).
Десяти- и пятнадцатидневные зародыши аксолота, освобожденные от яйцевых оболочек, легко живут в аквариумах и могут служить для опытов над воспалением. Возьмем кусочек ляписа и дотронемся им до края плавника предварительно кураризированного зародыша. Мы произведем очень ограниченный обжог; смоем затем избыток ляписа струей соленой воды. Вместо обжога можно произвести легкое повреждение плавника уколом иглы, предварительно погруженной в порошок кармина или индиго.
Оба эти способа убивают известное число клеток и обнажают часть плавника, которая разбухает от действия воды. Это действие передается также соседним клеткам; повреждение их выражается в том, что содержимое, особенно
звездчатых клеток, становится менее преломляющим свет и наполняется вакуолями. Через короткое время после операции известное количество блуждающих клеток направляется к пораненному месту. Эпидермис же стягивается, закрывая рану. Уже на другой день на месте повреждения наблюдается значительное, хотя и не обильное, скопление блуждающих соединительнотканных клеток. Они поглощают остатки разрушенных клеток и порошок краски, проникший в рану (рис. 47). Некоторые из скопившихся клеток находятся на стадии кариокинетиче-
Рис. 47. Воспаленный участок зародыша аксолота.
323
ского деления. Явление это, однако, слишком редко, чтобы можно было им объяснить происхождение многих клеток на месте повреждения. Такая гипотеза, впрочем, становится лишней ввиду прямого наблюдения притока блуждающих клеток к оперированному месту. Звездчатые клетки, за которыми можно следить изо дня в день на одном и том же животном, играют при этом совершенно пассивную роль.
Происходящие в них кариокинетические деления ничем не отличаются от подобных же явлений в других пунктах плавника. Кровеносные сосуды также не играют никакой роли в явлениях, следующих за повреждением. Сосуды эти проходят в хвостовой
Рис. 48. Участок хвостового плавника личинки тритона через 1/4 часа после прижигания ляписом: 1 — 5 — неподвижные клетки; а, b — блуждающие клетки.
части в виде двух больших стволов, посылающих в плавник только тоненькие ветви, лишенные кровообращения.
Итак, у зародыша позвоночного реакционные явления производятся исключительно соединительнотканными подвижными клетками, без всякого вмешательства сосудов или белых кровяных шариков.
Аналогия описанного явления с реакцией после повреждения у медуз и личинок иглокожих — неоспорима. Как здесь, так и там происходит скопление соединительнотканных фагоцитов у места повреждения.
Подобные же явления наблюдаются у молодых личинок аксолота и тритона, хвостовой плавник которых снабжен кровеносными сосудами. Последние, однако, относительно настолько малы, что крупные лейкоциты с трудом могут проходить сквозь их стенки. К тому же движение их затруднено чрезвычайно большими красными кровяными шариками. Поэтому миг-
324
рация происходит только в очень редких случаях, и мы наблюдаем следующее странное явление. Сосуды, находящиеся по соседству с поранением, остаются совершенно пассивными (не замечается даже видимого расширения их); напротив, блуждающие соединительнотканные клетки притекают к оперированной области.
Ввиду важности воспалительной реакции без участия сосудов у позвоночных, я остановлюсь еще на описании воспаления у молодых личинок Triton taeniatus. Край хвостового плавника
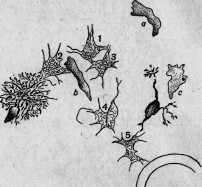
Рис. 49. То же через 3/4 часа после прижигания.
такой личинки был прижжен маленьким кристаллом ляписа и тотчас обмыт раствором поваренной соли и чистой водой. Поэтому повреждение распространилось на небольшое только число эпидермических и соединительнотканных клеток. Соседние звездчатые клетки стали менее преломляющими свет; они насытились жидкостью, вследствие чего в их протоплазме появились вакуоли и отростки их укоротились (рис. 48). Две из соседних блуждающих соединительнотканных клеток направились к поврежденному участку. В одной из близлежащих петель кровеносного сосуда кровообращение совершенно прекратилось. Через три четверти часа после операции количество блуждающих клеток несколько увеличилось в ближайшей области поранения. Все они переместились по направлению к прижженному месту (рис. 49).
Через три часа после начала опыта звездчатые клетки, сохранившие свое прежнее положение, получили вновь нормальную преломляемость. Они только обнаруживали некоторое изменение формы своих мельчайших разветвлений.
325
Число блуждающих клеток еще более увеличилось, но исключительно за счет соседней соединительной ткани. Кровообращение сосудов не изменилось, и в них не видно было никаких следов диапедеза. Через 5 часов после прижигания струп, образованный омертвелым эпидермисом, отпал и обнаружил новый эпидермический слой. Под ним наблюдалось скопление известного количества блуждающих клеток (рис. 50). Другие под-
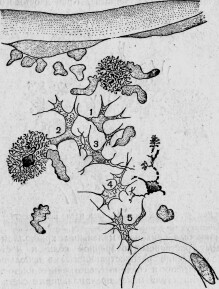
Рис. 50. Тот же участок через 5 часов после прижигания: 1 — 5 — те же неподвижные клетки, как на рис. 48, 49.
вижные клетки передвигались в том же направлении, как и в начале опыта, неподвижные же оставались неизменными. Кровообращение петли соседнего сосуда возобновилось, но диапедеза не обнаружилось. На следующий день поврежденная область вполне зажила. Звездчатые клетки (рис. 51) вернулись к своему нормальному состоянию, и отростки их вновь приняли форму оленьего рога. Блуждающие клетки в соединительной ткани были так же немногочисленны, как в нормальном состоянии. Часть их оставалась скопленной под возобновленным эпидермисом.
326
Ввиду такого быстрого заживления, прижигание было повторено на том же месте, но только сильнее. Неподвижные клетки обнаружили тотчас те же изменения, как и в первый раз; тело этих клеток разбухло, стало менее преломляющим, и в нем появились вакуоли; отростки же значительно укоротились. Кровообращение нескольких сосудистых петель прекратилось. Несмотря на силу повреждения, диапедез обнаружился только в самой незначительной степени. В течение пятичасового наблюдения я проследил выхождение одного только лейкоцита сквозь стенку сосуда. Это, однако, не помешало скоплению блуждающих клеток, часть которых направилась к поврежденному месту.
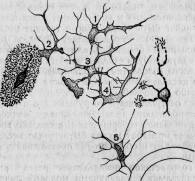
Рис. 51. Тот же участок через 24 часа после прижигания: 1 — 5 — те же фиксированные клетки, как на рис. 48—50.
Подобные наблюдения, повторяемые в течение нескольких лет и дававшие всегда тот же результат, ясно доказали возможность воспаления у позвоночных без вмешательства сосудистой и нервной систем. Это явление, чисто клеточного свойства, можно поставить рядом с воспалительной реакцией кольчатых червей.
Итак, генеалогическая картина воспаления, выясненная из наблюдений реактивных явлений у беспозвоночных, может быть дополнена исследованиями зародышей и молодых личиночных состояний позвоночных. Добытые факты доказывают, что первоначально реакция организма против повреждений была по существу сходна у обеих групп животного царства. Но в то время как у беспозвоночных она осталась неизменной, у позвоночных реакция эта заменилась гораздо более сложными явлениями. Уже у взрослых личинок тритона и аксолота — с
327
большим количеством кровеносных сосудов, достаточно крупных, чтобы дать свободный проход лейкоцитам, — воспалительная реакция происходит классическим путем.
Вследствие описанных повреждений кровообращение сначала ускоряется, а затем замедляется. Лейкоциты принимают периферическое положение в сосудах и затем мигрируют из них к месту повреждения.
Эти явления всего удобнее изучать на головастиках различных батрахий. У некоторых из них (как у Bombinator igneus) значительная величина и прозрачность плавников позволяют подробное наблюдение. Вызвав воспаление прижиганием маленьким куском ляписа, простым уколом или каким бы то ни было другим повреждением, можно на живом головастике проследить всю серию воспалительных и восстановительных явлений. Исследование это может быть продолжено в течение нескольких дней или недель. Такой способ представляет большое преимущество перед исследованием брыжейки или языка лягушки. Чтобы фиксировать какую-нибудь стадию, прибегают к методу Ранвье*: погружают целого головастика в алкоголь, разбавленный на 2/3 водою. После нескольких часов пребывания [головастика] в этой жидкости можно кисточкой снять весь слой эпидермиса. Тогда отделяют хвост от туловища и погружают его в ту же жидкость, к которой прибавляют несколько капель водного раствора везувина. Окрашивание совершается в несколько минут, так что промытый водою препарат может быть тотчас исследован в ней же. Это избавляет от необходимости обезвоживать его и заключать в бальзам.
Плавник головастиков представляет резкие анатомические отличия сравнительно с плавниками молодых личинок тритона и аксолота.
Кровеносные сосуды первых гораздо многочисленнее и богаче разветвлениями. Наоборот, в нормальной соединительной ткани головастика гораздо меньше блуждающих клеток. Кроме того, все гистологические элементы — как соединительнотканные клетки, так белые и красные кровяные шарики — меньших размеров. Соединение всех этих особенностей у головастика представляет удобные условия для миграции лейкоцитов. И действительно, последняя очень легко вызывается различными повреждениями. Мне удавалось вызвать ее как отрезыванием кончика хвоста, так и введением острого постороннего тела или прижиганием ляписом. Воспалительные явления обнаруживаются иногда уже через четверть часа после операции. При этом наблюдается расширение сосудов и значительная миграция клеток. В случае продолжительного раздражения, как, например, при введении занозы, воспалительную миграцию можно наблюдать в течение нескольких дней подряд. Она приводит к гораздо большему
* Traite technique d'histologie, изд. 2, 1889. 328
скоплению лейкоцитов у поврежденного места, чем при тех же условиях у молодых личинок тритона или аксолота. Сравнение реакционных явлений у хвостатых амфибий и у головастиков бесхвостых ясно показывает, что реакция эта значительно усиливается от вмешательства кровеносной системы.
Очень вероятно, что соединительнотканные блуждающие клетки также направляются к поврежденному месту. Но так как число их очень ограничено, то роль этих элементов совершенно сглаживается рядом с массой мигрировавших лейкоцитов. Неподвижные соединительнотканные клетки обнаруживают те же изменения, как и у тритона. Под влиянием раздражения они тотчас разбухают, делаются менее светопреломляющими, наполняются вакуолями, отростки их укорачиваются и теряют разветвления. Но вскоре эти клетки вполне восстановляются, сохраняя свою неподвижность и остальные характерные свойства.
Блуждающие клетки молодых хвостатых и лейкоциты головастиков и более взрослых личинок хвостатых, скопленные у места повреждения, тотчас обнаруживают свои фагоцитные свойства. Скопившиеся фагоциты эти жадно поглощают введенные с более крупными телами или просто втертые в рану зерна краски, а также остатки умерщвленных пигментных и других клеток. В своей работе о воспалении амфибии (Biolog. Centralbl., 1883) я настаивал и на том факте, что неподвижные звездчатые клетки соединительной ткани также обладают фагоцитными свойствами. После нескольких дней воспалительного процесса в плавнике в нем легко можно видеть клетки, заключающие в своей протоплазме посторонние тела, как, например, зерна кармина или остатки красных кровяных шариков. Клетки эти снабжены отростками, разветвленными наподобие оленьих рогов, и сходны с неподвижными, звездчатыми элементами. Из этих фактов, подтвержденных мною в несколько приемов, я вывел, что неподвижные клетки — такие же фагоциты, как и блуждающие. Но с тех пор мне пришлось убедиться в неверности этого объяснения. Несмотря на многочисленные попытки, мне никогда не удавалось констатировать захватывания посторонних тел протоплазматическими отростками неподвижных клеток. Исследования, предпринятые в этом направлении и повторенные в течение нескольких лет, убедили меня в том, что готовые, неподвижные клетки никогда не поглощают карминовых зерен или других посторонних тел. Если эти последние оказываются в них, то они были поглощены во время более ранних стадий, когда неподвижные клетки были еще подвижными фагоцитами. Такие факты служат прямым доказательством перехода блуждающих клеток в неподвижные элементы. Хотя вывод этот находится в противоречии с почти единогласным мнением всех патологов, тем не менее он вполне реален 170. Не следует, однако, заключать, что все скопившиеся на месте повреждения блуждающие клетки превращаются в неподвижные, звездчатые элементы.
329
Большое число фагоцитов не изменяется таким образом. Многие из них погибают и поедаются другими фагоцитами, как это всегда видно несколько дней после начала воспаления (рис. 52). Кроме того, некоторые подвижные клетки проникают в эпидермис, откуда они освобождаются наружу и погибают в окружающей воде; наконец, другие проникают в лимфатические сосуды и увлекаются потоком лимфы.
Несмотря на полное восстановление, некоторое количество фагоцитов остается все-таки на месте повреждения. В тех случаях, когда раздражитель продолжает оставаться в ткани, например стеклянные трубочки, введенные в соединительную ткань головастиков, некоторое количество мигрирующих лейкоцитов остается в поврежденном месте, окружая постороннее тело иногда в продолжение целых месяцев. Эта картина воспаления у головастика может служить примером того же самого процесса у других позвоночных, не исключая млекопитающих и даже человека. Мы подошли теперь к последней степени усложнения воспалительной реакции в животном царстве. Прежде чем рассматривать в подробности этот патологический процесс, необходимо бросить взгляд на эволюцию этого важного явления.
Главная причина воспаления, инфекция, должна быть рассматриваема как борьба двух организованных существ — паразита и его хозяина. Так как эта борьба приводит к приспособлению обеих сторон, то можно допустить, что организм вырабатывает средства для защиты против нападающих. Если одноклеточные организмы, у которых характер борьбы более очевиден, обладают уже средствами защиты, то невозможно допустить, чтобы высшие организмы были их лишены.
Растения, организмы пассивные, защищаются выделением толстых, устойчивых оболочек. Чтобы проникнуть через них, паразит должен принять особенные меры, т. е. выделить ферменты, растворяющие клетчатку, или силой роста оболочку пробуравить. Растения поэтому хорошо защищены против многих паразитов, неспособных к этому.
Паразиты, проникшие в растительную клетку, или другие какие-нибудь вредные деятели причиняют ей смерть. Но если она сама неспособна к возрождению, то другие, незараженные
330
Рис. 52. Фагоциты, содержащие в себе другие фагоциты. Плавник головастика Bombinator.


клетки или даже возбужденные раздражающей причиной, размножаются и пополняют недостаток, произведенный смертью непосредственно затронутых клеток.
Итак, в растительном царстве есть первичное повреждение и некроз, есть также восстановление, часто даже сверх нормального, но совсем нет воспаления.
Исключение из этого правила представляют некоторые низшие растения, именно пласмодии миксомицетов, так как они способны к передвижению и внутриклеточному пищеварению. Это последнее свойство, проявляющееся в захватывании и переваривании или выбрасывании вредного деятеля, содействует защите организма. Если оно само по себе не составляет еще воспалительной реакции, то по крайней мере представляет ее первоначальный источник.
Итак, воспаление появляется только в животном царстве и медленно развивается, начиная с существ, имеющих мезодерму. Вначале оно ничем не отличается от простого внутриклеточного пищеварения посредством подвижных мезодермических фагоцитов. Так, у губок пищеварительная и воспалительная функция еще соединены вместе. Но вслед за окончательным отделением энтодермы от мезодермы обе эти функции разделяются. Энтодерма функционирует исключительно как орган пищеварения, а мезодерма исполняет роль защиты против вредных деятелей, по возможности переваривая их. Мезодермические фагоциты сохраняют свойство внутриклеточного пищеварения и проявляют его, сливаясь в пласмодии, или просто соединяются, образуя капсулы вокруг паразитов и других посторонних тел. Фагоцитная реакция обнаруживается всеми мезодермическими фагоцитами. Ими бывают главным образом клетки соединительной ткани, перитонеальные элементы и клетки перивисцеральной жидкости или крови. Во всех случаях фагоциты борются с нападающим, захватывая его и переваривая.
Совершенно ясно, что воспаление позвоночных, где фагоциты, выходя из сосудистой системы, направляются к нападающему, отличается только количественно от подобных же явлений у беспозвоночных. Вследствие этого оно должно быть рассматриваемо как реакция организма против вредного деятеля. Итак, существенный источник, «primum movens», воспаления состоит в фагоцитной реакции животного организма. Все остальное составляет аксесуары этого явления и выражается в средствах облегчить приток фагоцитов к поврежденному месту 171.
Явления, приводящие к смерти, как, например, повреждение или первичный некроз, так же как и восстановительные явления, наступающие за воспалением, не относятся к этому последнему и не должны быть смешиваемы с ним.
С точки зрения сущности вопроса совершенно безразлично, с какой именно стадии развития фагоцитной реакции приписывается этой последней название воспаления. Можно ограничить этот
331
термин явлениями, сопровождающимися содействием сосудов, как этого требуют Штриккер*, Розер** и др. В таком случае фагоцитная реакция более взрослых личинок Urodelae войдет в рамку воспаления, тогда как та же реакция только более молодых личинок, где фагоциты доставляются соединительной тканью, не будет называться воспалительной. Или, если придерживаться этимологического значения слова, термин «воспаление» можно было бы применять в тех случаях, когда фагоцитная реакция сопровождается жаром, т. е. его можно бы было применять только к теплокровным животным. Самое важное будет заключаться, как и всегда, в определении естественных отношений явлений и в восстановлении их генеалогического развития.
Общее заключение, к которому привел нас сравнительный обзор фагоцитной реакции, облегчит более подробное изучение воспаления позвоночных.
* Allgemeine Pathologie der Infectionskrankheiten, Wien, 1886, стр. 112. ** Entzundung und Heilung, Leipzig, 1886, стр. 58.
ЛЕКЦИЯ ДВЕНАДЦАТАЯ
Приложение добытых фактов к критике теорий притяжения питательных веществ и повреждения сосудистых стенок. — Опыты Конгейма с языком лягушки. — Возбудители воспаления, введенные в кровь. — Реакции у беспозвоночных как аргумент против теории Конгейма. — Борьба организма против внешних деятелей. — Роль внутриклеточного пищеварения. — Фагоциты. — Hemitis. — Примеры: возвратный тиф, болезнь дафний. — Туберкулез. — Сущность воспаления. — Чувствительность фагоцитов. — Ее прогрессивный ход. — Чувствительность эндотелиальных клеток. — Определение воспаления. — Воспаление не есть регенерация. — Воспаление не есть резорбирование. — Возражения, сделанные против биологической теории воспаления. — Витализм. — Телеология. — Отсутствие фагоцитов в угрожаемых местах. — Несовершенство воспалительной реакции. — Активное вмешательство человека. — Сравнительная патология
Сделавши обзор главных явлений воспалительной реакции в животном царстве, мы можем спросить себя, насколько факты этой реакции согласуются с различными теориями, приведенными в первой главе.
Бесполезно долго останавливаться на невозможности объяснить эти факты теорией питания, требующей значительного притока питательных веществ к воспаленному месту, и ненормального размножения местных элементов. По мнению Вирхова*, воспаление начинается с того момента, когда появляется расстройство в питании. Расстройства состоят в способности «притягивать, т. е. непосредственно поглощать и изменять, смотря по обстоятельствам, большие количества питательных веществ». Здесь происходит усиленное питание клеток воспаленного органа на счет жидкой части крови. Смотря на вещи с этой точки зрения, Вирхов объясняет самые характерные воспалительные явления не как спасительную реакцию, но как процесс, существенная сторона которого проявляется в его остром характере и особенно в опасности его для организма (I. с., 399).
По теории Самуэля и Конгейма, сущность воспаления состоит в молекулярном повреждении сосудистой стенки. Измененная вследствие какого-нибудь вредного влияния, стенка теряет свою способность удерживать элементы крови, которые и . проходят совершенно пассивно наружу, чтобы направиться к месту наименьшего сопротивления. Воспаление не есть, следовательно, реакция организма против внешних деятелей, но просто первичное
* Cellularpathologie, изд. 4, 1871, стр. 475.
333
повреждение кровеносных сосудов. Один опыт Конгейма очень резко выясняет его точку зрения. Если с помощью лигатуры остановить на 48 часов кровообращение в языке лягушки и если потом расправить язык, то оно восстановляется, но со всеми признаками воспалительного кровообращения, т. е. с периферическим расположением лейкоцитов, сопровождаемым диапедезом. Этот факт, по мнению Конгейма, есть прямое следствие повреждения сосудистой стенки от продолжительной анемии. «Я признаю неоспоримым, — прибавляет Конгейм*, — что причину воспаления в этом случае надо искать в самих сосудах; все, что происходит вне сосудов, производит впечатление слишком второстепенных изменений, чтобы ими можно было что-нибудь объяснить». Между тем вне сосудов происходят очень важные явления. Периферические ткани, лишенные питания и защиты крови, должны подвергнуться нападению со стороны микробов, находящихся в изобилии в ротовой полости. Все ткани или некоторые из них должны в то же время претерпевать явления вырождения, так что все это может дать внешнее возбуждение, достаточное для того, чтобы вызвать воспалительную реакцию. Пример, приведенный Конгеймом, вполне выясняет его теорию, но не доказывает ее.
Чтобы вызвать центральное, так сказать, повреждение сосудов, не зависимое от повреждения других органов, расположенных на периферии, есть очень простое средство, а именно — вверти раздражителя внутрь самих сосудов. В той же работе Конгейма «об эмболиях» приводятся опыты, которые должны произвести эмболические абсцессы. С этой целью Конгейм впрыскивал в артерии лягушек множество веществ, которые должны были вызвать значительное воспаление, например, шарики ртути, порошок шпанской мухи, частицы сгнившей говядины, но все было напрасно. Никогда эти вещества не производили воспаление, тогда как, введенные под кожу, они вызвали очень сильную реакцию. Если воспаление есть только изменение сосудистых стенок, то как объяснить отсутствие этого изменения в случаях непосредственного влияния раздражающей причины?
Рядом с опытами Конгейма можно привести много других фактов, дающих тот же результат. Часто патогенные микробы, обыкновенно столь сильные возбудители воспаления, не производят никаких экссудативных явлений, если они находятся в крови. В возвратном тифе кровь переполнена спириллами, которые своими винтообразными движениями, а также, вероятно, своими токсинами, действуют прямо на сосудистую стенку.
По теории Конгейма, эти стенки должны были бы неизбежно быть изменены и дать проход элементам крови наружу. Между тем при возвратном тифе, когда тело больного «воспалено» в высшей степени, в органах вовсе не происходит воспаления. Многие другие микробы, например сибиреязвенные, производят
* Die embolischen Processe, 1873, стр. 51. 334
сильное воспаление, если их прививать под кожу, и не причиняют ровно никакого воспаления, в смысле Конгейма, если они находятся в крови.
Подобные же явления наблюдаются при впрыскивании туберкулезных бацилл в кровь, между тем как при введении их вне сосудов неизбежно происходит экссудативное воспаление.
Кроме этих фактов, вовсе не объясняемых теорией первоначального повреждения сосудистых стенок, вся сравнительная патология говорит против взгляда Конгейма. Реакционные явления у беспозвоночных показывают, что инфильтрация предшествует сосудистым явлениям и что лейкоциты не профильтровываются пассивно, а направляются к поврежденному месту с помощью своей чувствительности и амебовидных движений.
Но если теории Вирхова и Конгейма не могут быть приняты при настоящем состоянии наших знаний, нужно ли поэтому оставлять всякую надежду понять воспалительные явления и довольствоваться их простым описанием, как это делает большая часть современных патологов?
Изучение воспаления с точки зрения сравнительной патологии показывает прежде всего, что это явление есть по существу реакционное. Организм, угрожаемый каким-нибудь вредным деятелем, защищается средствами, которыми он располагает. Так как мы видели, что даже самые низшие одноклеточные животные не относятся пассивно к вредным для них деятелям, а, напротив, борются с ними, то неужели же наиболее развитые организмы, как человек и млекопитающие, не будут действовать так же? Итак, должна быть неоспоримая борьба зараженного организма против вредного деятеля; но в чем же состоит эта борьба? Эволюция воспаления доказывает, что именно это явление есть наиболее распространенное и наиболее деятельное средство защиты в животном мире.
«Primum movens» воспалительной реакции есть пищеварительное действие протоплазмы, проявляемое относительно вредного деятеля. Это действие, свойственное всему или почти всему организму простейших, принадлежит всей пласмодической массе миксомицетов, а начиная с губок сосредоточивается в мезодерме. Фагоцитные клетки этого слоя приближаются, захватывают и разрушают вредного деятеля в тех случаях, когда зараженный организм остается победителем. Фагоцитная реакция бывает сначала медленной, так как единственным средством фагоцитов приблизиться к вредному деятелю служат их амебовидные движения; с появлением же кровеносной и сосудистой систем реакция значительно ускоряется. С помощью кровяного потока организм в каждую данную минуту может посылать к зараженному месту значительное число фагоцитов. В тех случаях, когда кровообращение совершается в системе лакун, приток фагоцитов происходит без специальных приспособлений. Когда же эти клетки заключены в закрытых сосу-
335
дах, то для достижения своей цели — защиты организма — им необходимо особое приспособление, именно диапедез.
Допустив, что воспаление высших животных есть спасительная реакция организма и что диапедез — часть этой реакции, мы увидим, что воспалительные явления становятся простыми и ясными. Уже давно были поражены наблюдением лопастей и полиморфной формой ядра гнойных клеток. Эта особенная форма свойственна многоядерным лейкоцитам, составляющим большую часть всех белых кровяных шариков (75 : 100).
Наблюдая, что большое количество клеток гноя погибает в экссудате, связали этот факт со странной формой ядра. Утверждали и продолжают утверждать еще до сих пор, что многоядерные лейкоциты суть клетки, предназначенные к смерти, не способные к значительной деятельности. Но именно лейкоциты и являются наиболее деятельными клетками организма. Форма же их ядра гораздо лучше объясняется специальным приспособлением к прохождению сквозь сосудистые стенки. При наблюдении за диапедезом бросается в глаза трудность, с какой сопряжен проход ядра. Если последнее находится уже вне сосуда, то протоплазма переходит стенки почти мгновенно. Понятно, что лопастное ядро должно гораздо легче проходить стенку, чем большое цельное ядро. Вследствие этого в гное находится гораздо больше многоядерных лейкоцитов, чем одноядерных; потому же многоядерная форма ядра совершенно отсутствует у беспозвоночных (исключая некоторых головоногих) и находится только у лейкоцитов, приспособленных к диапедезу.
Факты, доказавшие невозможность принять теорию Конгейма, легко могут быть объяснены с точки зрения защищаемой здесь нами теории. Если раздражающий деятель находится вне сосудов, он вызывает типическое воспаление, сопровождаемое диапедезом. Если тот же самый деятель находится внутри сосудов, диапедеза вовсе не происходит, но лейкоциты борются против микроба в самой крови. Возьмем пример возвратного тифа, где спириллы, очевидно, действуют на стенку сосудов, не вызывая диапедеза. Но лейкоциты увеличиваются в числе, и происходит лейкоцитоз, ведущий к борьбе, конец которой характеризуется захватыванием спирилл лейкоцитами. Мы имеем здесь случай воспаления, не сопровождаемый диапедезом; борьба фагоцитов со спириллами совершается в самой крови. Несмотря на отсутствие воспалительного диапедеза, возвратный тиф сопровождается сильным жаром и другими явлениями, указывающими, что мы имеем дело с воспалительной болезнью. Это будет, таким образом, пример воспаления самой крови, род «hemitis», как думал прежде Пиорри.
Это есть общее явление для всех случаев борьбы у тех животных, кровеносная система и общая полость тела которых соединены вместе. Так, в болезни дафний (производимой Monospora) часто происходит значительное скопление лейко-
336
цитов вокруг спор, и это скопление бывает именно в кровеносной полости.
Возьмем еще пример туберкулеза. При впрыскивании под кожу туберкулезные бациллы производят воспаление, сопровождающееся значительным диапедезом. При впрыскивании тех же бацилл прямо в кровь диапедеза не происходит, но фагоциты соединяются вокруг бацилл внутри сосудов и образуют внутри-сосудистые туберкулы. Нельзя было бы сказать, что в первом случае (внесосудистое впрыскивание) воспаление происходит, а во втором (внутрисосудистое впрыскивание) оно отсутствует. Тем более нельзя этого сказать, что в обоих случаях происходят одинаковые туберкулы. Вот еще пример воспаления самой крови.
Все эти случаи внутрисосудистого воспаления без диапедеза, так же как и воспалительные явления у молодых личинок аксолотов и тритонов (где мигрирующие клетки соединяются вокруг поврежденного места), наконец, целый ряд реакционных проявлений у стольких беспозвоночных ясно доказывают нам, что существенный и первичный элемент типичного воспаления состоит в реакции фагоцитов против вредных деятелей. Если этот последний находится в общей полости, наполненной кровью, фагоциты соединяются в общей полости; если этот фактор входит внутрь сосудов, как в возвратном тифе или при внутрисосудистом. туберкулезе, то соединение фагоцитов происходит в самой крови; если, наоборот, вредный фактор находится вне кровеносной полости или вне сосудов, то происходит миграция фагоцитов к угрожаемому месту, и при этом миграция без диапедеза (беспозвоночные, молодые личинки хвостатых амфибий) или с диапедезом (позвоночные).
Для появления фагоцитной реакции надо прежде всего, чтобы клетки были возбуждены положительным образом. Отрицательная чувствительность также может служить средством защиты для подвижного организма, как, например, пласмодию миксомицетов, который удаляется от вредного фактора. В случаях проникания этого последнего внутрь организма отрицательная химиотаксия фагоцитов оставляет паразиту поле сражения, что ведет за собой смерть зараженного организма, как это часто и случается. Вот почему в ряду этих существ мы видели прогрессивную эволюцию положительной чувствительности лейкоцитов. У дафний поражает множество болезней, в которых фагоцитоз вполне или почти вполне отсутствует. У амфибий положительная химиотаксия уже очень ясно выражена, но у кролика, как доказал Габричевский, она гораздо сильнее. У грызунов и вообще у мелких лабораторных животных наблюдается много смертельных болезней, как, например, куриная холера, вибрионная септицемия птиц, при которых фагоцитоз часто вполне отсутствует. У человека и высших млекопитающих подобные болезни несравненно менее часты.
337
Но рядом с подвижными фагоцитами, направляемыми своей чувствительностью, есть еще неподвижные, развитые особенно у позвоночных и представителями которых являются эндотелиальные клетки сосудов. Так как эти клетки сократимы и обладают фагоцитными свойствами, то естественно предположить, что они также и чувствительны. Замечательное явление взаимного притяжения протоплазмических придатков капилляров при их развитии и образовании вследствие этого сосудистой петли может быть легко объяснимо химиотаксией эндотелиальных элементов. То же самое объяснение может служить для тех случаев, где (как во многих новообразованиях, в глазном panus'e и т. д.) сосуды легко проникают и обильно развиваются в зараженной ткани. Это разрастание сосудов может происходить вследствие химиотаксии или другой какой-нибудь положительной чувствительности, тогда как их отсутствие в гранулемах, как туберкулез, лепра, актиномикоз, могло бы быть объяснено отрицательной чувствительностью. Содействие эндотелиальных клеток в воспалительном процессе, поскольку оно происходит вследствие их сократительности, также объясняется чувствительностью этих элементов.
Наконец, является еще иная чувствительность, именно чувствительность нервных элементов, которые присоединяются к фагоцитному и сосудистому аппаратам, чтобы облегчить реакцию против вредных деятелей.
Итак, воспаление в своем целом должно быть рассматриваемо как фагоцитная реакция организма против раздражающих деятелей; эта реакция выполняется то одними подвижными фагоцитами, то с содействием сосудистых фагоцитов или нервной системы.
Теория, изложенная в этих нескольких строках, может быть названа биологической или сравнительно-патологической теорией воспаления, так как она основана на изучении явлений жизни клеток, наблюдаемой с точки зрения сравнительной патологии.
Прежде всего необходимо подчеркнуть тот факт, что существенные явления воспаления представляют действительно борьбу фагоцитов против раздражающего деятеля. Мы видели, что лейкоциты могут превращаться в неподвижные клетки, и можно думать, что их значительные скопления совершаются именно с этой целью. Предположение это должно быть, однако, оставлено вследствие того факта, что у высших позвоночных многоядерные лейкоциты, играя роль при воспалении, вовсе не содействуют образованию грануляций. В развитии этих последних принимают деятельное участие, рядом с эндотелиальными и соединительнотканными клетками, только одноядерные лейкоциты. Эти лейкоциты образуются прямо за счет многочисленных лимфоцитов, приходящих к местам регенерации.
Но, может быть, лейкоциты, соединенные в воспалительных очагах, служат только к поглощению мертвых клеток и мерт-
338
вых микробов? Их часто называли простыми «подметателями» организма. Мы уже видели, что это предположение совсем не подтверждается и что лейкоциты захватывают живых паразитов, к которым направляются с самого начала инфекции. Эти доказательства, хотя и прямые, могут показаться недостаточными, но их можно подтвердить еще более.
Если сущность лейкоцитной реакции при воспалении сводится к поглощению твердых частей, то вообще случаи обильных и быстрых поглощений должны были бы иметь очень ясно выраженный характер воспаления. Но этого нет. Метаморфоз лягушек сопровождается поглощением личиночных органов — хвоста и жабр, — что происходит очень быстро, в несколько дней, с помощью фагоцитов, захватывающих все ткани в свою протоплазму. А между тем в данном случае совсем не происходит воспаление, и для поглощения этих элементов достаточно бывает фагоцитов самих тканей. Поглощение, очевидно, гораздо легче, чем борьба с паразитами, и требует гораздо менее значительной фагоцитной деятельности.
У млекопитающих настоящими «подметателями», т. е. резорбирующими фагоцитами, являются макрофаги в общем и одноядерные лейкоциты в частности. Эти клетки играют важную роль, особенно в хронических воспалениях, как, например, туберкулез. При острых же инфекциях, наоборот, микрофаги, или нейтрофильные многоядерные лейкоциты, принимают участие в борьбе. При роже, например, одни только многоядерные лейкоциты, а не макрофаги, захватывают стрептококков. Все же резорбирование выполняется, наоборот, макрофагами, и особенно часто они захватывают микрофагов, многие из которых погибают в борьбе и должны быть сами резорбированы. Если бы воспалительная миграция имела исключительной функцией резорбирование, то образование гноя ее при окончании было бы бессмысленным, так как гной есть масса лейкоцитов, из которых большое количество умирает и в свою очередь должно быть резорбировано. Вполне естественно, напротив, предположить, что миграция есть реакция организма в борьбе, во время которой множество главных борцов, микрофагов, погибает на поле битвы. Резорбирование наступает всегда за этим и выполняется другим родом фагоцитов.
При первом моем изложении биологической теории воспаления, восемь лет тому назад*, я высказал мысль, что эта реакция выполняется с помощью живой связи «между соединительнотканными клетками, элементами эндотелия и лейкоцитами, которые все вместе образуют как бы живую цепь, выполняющую главную роль в воспалении у позвоночных». Соединительнотканные клетки, затронутые прежде всего, передают действие сосудистой стенке, клетки которой сокращаются и облегчают прохождение белым кровяным шарикам. В то время имели право
* Biologisches Centralblatt, 1883, стр. 564.
339
предполагать только осязательную чувствительность различных клеточных элементов, хотя некоторые факты, как миграция на большие расстояния половых клеток гидрополипов*, давали уже возможность предсказывать существование химиотаксии. Впоследствии химиотаксия была доказана самым положительным образом.
Эту биологическую теорию часто находили слишком виталистической. Как пример наиболее искренно высказанного мнения, я приведу слова Френкеля**: «Теория фагоцитов, — говорит он,— предполагает удивительные свойства протоплазмы лейкоцитов; ей приписывают почти настоящие чувства, мысли и действия, род психической деятельности». Чувствительность фагоцитов вовсе не есть гипотеза, которую можно произвольно принять или отбросить; это есть прочно установленный факт, который нельзя игнорировать, как это делает Френкель. Что же касается упрека, сделанного мне Френкелем в том, что я приписываю лейкоцитам способность мыслить и хотеть, то ясно, что об этом нет и речи. Если чувствительность лейкоцитов, так же как и различных одноклеточных растительных и животных организмов, представляет первый шаг в длинном ряду явлений, приводящих к психической деятельности, то это совсем не удивительно. Психические явления не имеют чего-нибудь строго специфического и развиваются из усложнений очень простых актов, представляемых нам низшими организмами и клетками различных животных. Ученые, изучавшие ближайшим образом эти вопросы, как Спенсер, Ромэнс и др., хорошо это знают. Витализм и одухотворение, несправедливо приписываемые теории фагоцитов, должны быть поставлены в упрек скорее моим противникам, утверждающим, что психические акты самих животных представляют нечто совсем отличное от простейших явлений, свойственных низшим организмам.
Совершенно ошибочно также приписывают телеологический характер фагоцитной теории, рассматривающей воспаление как реакцию организма против раздражающих деятелей. Вся эта теория основана на законе эволюции, по которой свойства, полезные организму, сохраняются естественным подбором, тогда как вредные постепенно уничтожаются. Из низших животных выживают те, у которых подвижные клетки, выходя на борьбу с врагом, захватывают его и разрушают; другие же, фагоциты которых не функционировали, должны необходимо погибнуть. Вследствие подобного естественного подбора полезные свойства и в числе их те, которые служат для воспалительной реакции, устанавливались и передавались без заранее предустановленной какой-нибудь цели, как это должно бы было делаться с телеологической точки зрения.
* См. Weissmann. Die Entstehung der Sexualzellen bei Hydromedusen, Jena, 1883.
** Grundriss der Bacterienkunde, Berlin, 1890, изд. 3, стр. 203. 340
Много раз делали также следующее возражение*: если фагоцитная реакция развилась для защиты организма против угрожаемой опасности, то как объяснить, что именно в тех случаях, когда организм находится в особенной опасности, фагоциты отказываются выполнять свою роль. Это возражение есть также следствие неполного знакомства с принципами теории. Именно потому, что защита фагоцитов развивается по закону естественного подбора, а не вследствие предначертанной заранее цели, вполне понятно, что бывают случаи, где фагоциты не исполняют своей роли, что угрожает организму большой опасностью и смертью. В природе встречаются различные признаки, то полезные, то вредные организму. Первые способствуют выживанию, вторые — смерти. Возьмем, например, два организма: один, фагоциты которого легко отталкиваются микробами, и другой, у которого, наоборот, фагоциты обнаруживают положительную чувствительность, обусловливающую значительный фагоцитоз. Первый скоро станет добычей паразитов, будет уничтожен естественным подбором, тогда как второй будет сопротивляться инфекции, выживет и произведет потомство, одаренное теми же фагоцитными свойствами. Очевидно, что при таких условиях деятельность фагоцитов, поддерживаемая подбором, усилится.
Но целительная сила природы, главный элемент которой составляют воспалительные реакции, вовсе не есть еще приспособление, достигшее совершенства. Частые болезни и случаи преждевременной смерти достаточно это доказывают. Фагоцитный аппарат еще не достиг последней степени своего развития и находится на пути совершенствования. Очень часто фагоциты удаляются от неприятеля или разрушают элементы организма, часть которого они сами составляют (как при склерозах). Это несовершенство сделало необходимым деятельное вмешательство человека, не удовлетворенного функцией своей естественной целебной силы.
Защита организма против вредных деятелей, сконцентрированных в фагоцитном аппарате и соматической нервной системе, распространилась и на нервный аппарат психической деятельности. К нервным клеткам, управляющим сокращением и расширением сосудов, присоединяются клетки, производящие мысль и волевые акты. Результатом функций этих психических клеток явилась целая наука, имеющая целью защиту организма против вредных факторов.
Чтобы усилить целебное воспаление, наука изобрела разные методы (как во многих случаях искусственных повреждений), облегчающие воспалительную реакцию. Применение деятелей, возбуждающих воспаление, как жеквирити, бленоррейного яда, туберкулина и кантаридина, представляет сознательное продолже-
* См. Баумгартен, Berliner klin. Wochenscher., 1884, и Бердон-Сандерсон, British med. Journ., 1891, стр. 1085.
341
ние мер защиты, выработанных бессознательно целым рядом существ в их борьбе за существование.
Но как бессознательный реакционный аппарат — целебные силы природы с их фагоцитами — далеко не совершенен, так и сознательный реакционный аппарат — медицина — вовсе не представляет совершенства. Чтобы достигнуть своей конечной цели, она должна почерпнуть знания во всех сложных научных отраслях и, между прочим, в биологии, изучающей организмы и их эволюцию.
Применение сравнительно-патологической точки зрения полезно не только при изучении воспаления; другие задачи медицины тоже могут получить свою выгоду от этого сравнительного метода.
При изучении лихорадки надо было бы начать с исследования производства тепла низшими организмами и стараться подметить первые шаги этого производства у позвоночных, например у рептилий, чтобы уловить первоначальные проявления лихорадочной реакции. Так как многие из этих явлений развились в предшествовавшие геологические эпохи, то следовало бы еще составить понятие об условиях, в которых жили первые теплокровные животные.
В третьем важном патологическом вопросе — об опухолях — сравнительная патология может оказать неоспоримые услуги. Так как многие низшие организмы, растительные и животные, подвержены образованию опухолей, то гораздо легче на них констатировать роль паразитов в этиологии этих болезненных образований и отбросить теорию их происхождения из остатков зародышевых пластов.
Вопрос об атрофиях, тесно примыкающий к вопросу о хронических воспалениях, тоже должен рассматриваться с точки зрения сравнительной патологии. Атрофии представляют собой еще одну главу в патологии, в которой фагоцитарные явления играют вполне первенствующую роль.
Но если, с одной стороны, медицина может многое почерпнуть от биологии, часть которой она сама составляет, то и она может быть не бесполезна для последней. Общая биология, в свою очередь, может получить большую выгоду, присоединяя к предметам своего исследования болезненные явления, изучаемые патологией. Очень часто биология испытывает затруднения при изучении процессов эволюции, так как эти явления представляются ей уже в оконченной форме. Так, для ближайшего анализа актов естественного подбора явления, встречающиеся в природе слишком уравновешенными, вовсе не доставляют благоприятного материала. Чтобы выставить на вид действие этого общего закона, надо изучать менее стойкие явления, менее совершенные аппараты, одним словом, явления, в которых естественный подбор можно наблюдать каждый день. Таковы именно болезненные явления с вызываемыми ими реакциями, борьба между организмом и его
342
врагами; такие явления представляют лучший случай для изучения хода естественного подбора. В этой борьбе постоянно имеются выживающие, избранные подбором, и умирающие, исключенные тем же фактором. С одной стороны, избранными оказываются победители-организмы, а уничтоженными — паразиты; с другой стороны, исключаются побежденные организмы и избираются торжествующие паразиты.
Итак, я кончаю тем, чем начал. Общая патология должна быть соединена с зоологией или скорее с биологией, чтобы составить ее отрасль — сравнительную патологию. Развитие этой зарождающейся науки еще впереди, а между тем она уже теперь может оказать услуги медицине. Облегчая анализ реакционных явлений, она указывает элементы, которые должны быть особенно тщательно охраняемы в борьбе организма против своих врагов, и таким образом содействует решению одной из величайших задач человечества.