К истории учения о зародышевых листках. — Гипотезы о происхождении Metazoa. — Генеалогическое значение трех первых стадий дробления. — Образование энтодермы. — Замечания о теориях гастреи, планулы и плакулы. — Формулировка теории фагоцителлы. Ответ на некоторые возражения. — Родство фагоцителлы с Turbellaria Acoela. — Замечания по поводу теории щелевидного бластопора. — К генеалогии порошицы. — Генеалогические [соображения] об образовании мезодермы
Некоторые выводы, дающие первые принципы современной сравнительной эмбриологии, относятся еще к первой четверти нашего столетия. Так, натурфилософской школой в Германии было высказано положение: «Evolutionem, quam prima aetate quod subit animal, evolutioni, quam in animalium serie observandam putant, respondere»**.
Бэр *** высказался против этого вывода, когда он утверждал, что эмбриональные стадии животных не могут быть сравниваемы с другими взрослыми формами, но только с их эмбрионами. В конце концов, однако, он был вынужден и сам признать, что различие между двумя взглядами не столь уж велико, как это кажется сначала.
Значительный шаг вперед был сделан Л. Агассицем, когда он установил параллелизм между эмбриональными стадиями современных животных и главными ступенями геологической истории животного мира, но он не сумел из этой истины сделать правильных выводов.
* Седьмая, заключительная глава из большой работы И. И. Мечникова «Эмбриологические исследования над медузами», опубликованной на немецком языке в 1886 г. Полностью в русском переводе [Ю. И. Полянского] см. в «Избранных биологических произведениях», М., 1950, а также «Академическое собрание сочинений», т. 3, М., 1955 (перевод О. В. Воиновой). — Ред.
** Развитие, которое претерпевает с раннего возраста животное, соответствует развитию, которое полагают наблюдаемым в ряду животных (лат.). См. также прим. 33. — Ред.
*** «Ueber die Entwicklungsgeschichte der Tiere», т. I, 1828, схолий V, стр. 199 и далее111.
210
В то время как, с одной стороны, искали сходства между эмбриональными состояниями животных и взрослыми формами, а с другой — устанавливали такие же сходства эмбрионов с вымершими формами, Гексли в 1849 г. провел сравнение между зародышевыми листками позвоночных и главными слоями тела кишечнополостных, которые он назвал эктодермой и энтодермой.
Эта мысль в Англии не только не осталась незамеченной, но была популяризирована и дальше обобщена Гербертом Спенсером в одном из его прекрасных трактатов «О социальном организме»*. Английский философ писал: «Во всем животном царстве, начиная с Coelenterata**, первый фазис развития один и тот же. В зародыше полипа, равно как и в человеческом яйце, масса клеточек, из которых должно образоваться живое существо, отличается периферическим слоем клеточек, слегка различествующих от остальных клеток, которые этот слой окружают; впоследствии он разделяется на два слоя: внутренний, находящийся в прикосновении с желтком и называемый слизистым слоем, и наружный, подвергающийся окружающим влияниям и называемый серозным слоем, или, говоря словами профессора Гексли, употребляемыми им при описании процесса развития Hydrozoa, образуется ectoderm и entoderm***. Это первобытное разделение обозначает основное различие частей в будущем организме». В дальнейшем изложении следует установление аналогий эктодермы с высшими слоями человеческого общества, энтодермы — с низшими слоями и среднего листка — с третьим сословием 113.
В Германии учение Гексли долгое время не находило приверженцев, что было связано с известной реакцией, имевшей здесь место против общего применения учения о зародышевых листках и нашедшей свое наивысшее выражение в эмбриологии двукрылых Вейсмана 114. Подобное направление вполне соответствовало господствовавшей в то время теории типов, согласно которой все морфологические сравнения возможны лишь в пределах одного и того же типа животных.
В то время как в течение долгого периода при установлении сходств между животными в их анатомии и стадиях развития видели выражение общего плана, который понимали в чисто идеалистическом (ideale) смысле, в позднейшие десятилетия вместе с Дарвином узнали, что в основе этих сходств лежит генеалогическое родство. Дарвин особенно подчеркивал, что эмбрион, менее измененный, чем взрослое животное, позволяет судить о строении предков и что сходство зародышей указывает на общее происхождение взрослых стадий. Он старался эти
* Впервые было опубликовано в январе 1860 г. в Westminster Review. 3a отсутствием оригинала цитирую по русскому переводу122.
** Кишечнополостных (лат.). — Ред. *** Эктодерму и энтодерму (англ.). — Ред.
211
общие эмбриологические закономерности подтвердить конкретными примерами; при этом Дарвин объяснял наблюдавшиеся исключения изменениями хода развития, происходящими благодаря приспособлениям ранних стадий и их наследованию. Дарвин пришел к выводу, что установленный Агассицем параллелизм возник благодаря генеалогическому родству животных, что этот параллелизм проявляется только тогда, когда не происходит каких-либо затемняющих вторичных изменений в ходе развития.
В замечательной работе Фрица Мюллера «За Дарвина» эмбриологические принципы Дарвина были не только развиты дальше дедуктивным путем, но на отдельных конкретных примерах, взятых из эмбриологии и истории развития низших организмов, углублены и аналитически.
Вместе с тем им было показано, как следует обрабатывать сравнительно-эмбриологические данные. Ф. Мюллер настойчиво подчеркивает, что история развития только тогда повторяет генеалогический путь, когда потомки на пути к родительской форме не отклоняются рано или поздно, а проходят этот путь без уклонений, «но тогда, вместо того, чтобы остановиться, идут еще дальше». «В короткий промежуток времени нескольких недель или месяцев сменяющиеся формы эмбрионов и личинок дают нам более или менее верную картину тех изменений, благодаря которым в течение бесчисленных тысячелетий вид достиг своего настоящего состояния» («За Дарвина», стр. 76).
В соответствии с мыслями Дарвина об изменениях в ходе развития Ф. Мюллер выдвигает следующий тезис: «Историческое свидетельство, сохранившееся в истории развития, понемногу стирается вследствие того, что развитие принимает все более прямой путь от яйца к окончательной форме животного; оно часто искажается борьбой за существование, которую должны вести свободноплавающие личинки» (стр. 77)115.
После появления пролагающей новые пути работы Ф. Мюллера и отчасти под ее влиянием началась интенсивная разработка фактической стороны эмбриологии животных, причем низшие животные изучались в большей степени, чем высшие позвоночные 116. Совершенно независимо от этого направления Келликер в 1865 г. (во второй части своих «Icones histologicae», 90) пришел к выводам, которые в основном совпадают со взглядами Гексли. «Как бы то ни было, — пишет он в конце своего сообщения, — во всяком случае соответствие в строении гидроида и молодого зародыша позвоночных очень бросается в глаза, и, несомненно, дальнейшее изучение этого вопроса, сопровождающееся исследованием строения и гистологии развитии многих животных, приведет к нахождению простого закона развития». Эти дальнейшие исследования не заставили себя долго ждать. Несколько забытое в отношении беспозвоночных учение о зародышевых листках было вновь воспринято и обогащено
212
множеством новых фактов, так что скоро оно стало основой нового направления в эмбриологии. Важнейший толчок этому дало создавшее эпоху открытие Ковалевским* развития ланцетника: в эмбриологии этого животного были найдены явления, которые объединяли развитие позвоночных и беспозвоночных животных. После того как Ковалевский открыл двуслойную, покрытую ресничками личинку ланцетника, он начал искать аналогичные формы развития у других животных, причем ему удалось установить большое количество очень ценных фактов.
В то время как усилия этого рода исследователей сосредоточились вокруг нахождения основных стадий эмбрионального развитая, которые могли быть поставлены в параллель с явлениями развития у ланцетника и преимущественно относились к низшим животным с просто устроенными личинками, я обратился к разработке эмбриональных процессов высших беспозвоночных с намерением установить и у них учение о зародышевых листках. Сначала я исследовал эмбриологию Sepiola**, у которой нашел два зародышевых листка и проследил их участие в образовании органов. Среди членистоногих мне не удалось с достаточной ясностью обнаружить зародышевые листки у насекомых; зато я нашел их у высших ракообразных*** (Nebalia) и скорпиона****. У последнего я сначала (1866) нашел только два листка, после чего вскоре (1868) обнаружил и третий. Проведенные мною наблюдения показали, что у скорпиона верхний листок дает нервные клетки центральных органов, средний расщепляется на два листка с образованием сегментов, причем полость тела возникает путем соединения полостей отдельных сегментов и что, далее, нижний зародышевый листок оказывается
* История развития ланцетника (Amphioxus lanceolatus). Магистерская диссертация на русском языке в 1865 г. и Entwicklungsgeschichte des Amphioxus lanceolatus. Mem. Acad. Sciences de St.-Petersbourg [т. XI, № 4], 1867.
** История развития Sepiola. Диссертация на русском языке, 1867 г. Извлечение дано Клапарэдом в Archives des Sciences physiques et naturelles, 1867. Сравнить с появившейся в 1874 г., Zeitschr. f. wiss. Zool., т. XXIV, работой Гренахера о развитии головоногих, которая написана еще в прежнем направлении и в которой нет и речи о зародышевых листках.
*** История развития Nebalia. Диссертация на русском языке, 1868. **** Entwicklungsgeschichtliche Beitrage, 1868. Melanges biologiques de 1'Acad. Sciences de St.-Petersbourg. Против зародышевых листков у скорпиона очень решительно выступил Ганин в 1867 г. в своей русской диссертации об эмбриологии скорпиона и в Zeitschr. f. wiss. Zool,, т. XX, 1870, стр. 518. Когда Ковалевский (Embryol. Studien uber Wurmer u. Arthropoden, 1871, стр. 5) утверждает, что Ганин сделал «положительные наблюдения, что нервная система возникает из верхнего клеточного слоя», то это основано на недоразумении, потому что упомянутый исследователь выводит нервную систему скорпиона непосредственно из зародышевой полоски и не допускает дифференцировки на клеточные слои. Также неверно, когда Ковалевский приписывает Ганину открытие кишечно-железистого листка.
213
кишечно-железистым. Эти факты позволили мне в 1869 г. придти к заключению*, что «три листка зародыша скорпиона вполне соответствуют трем ремаковским листкам позвоночных животных». При этом меня не удерживало от этого вывода и то обстоятельство, что, как мне казалось, нервные волокна возникают не из верхнего, а из среднего листка. Подобный путь образования периферической нервной системы считался в то время правилом и для позвоночных животных 117.
Таким образом, для того чтобы получить достаточный фактический материал, сравнительно-эмбриологические исследования проводились с двух сторон. Лишь после того как я проследил в общих чертах зародышевые листки скорпиона, Ковалевский ** начал свои исследования по эмбриологии Oligochaete и насекомых, в процессе которых он обнаружил те же три листка и подробно проследил их судьбу. Обновленное учение о зародышевых листках у беспозвоночных получило прочный фундамент; возникло новое сравнительно-эмбриологическое направление, разрабатываемое главным образом немецкими и русскими исследователями 118.
Понятие зародышевых листков заимствовано из эмбриологии высших животных и перенесено затем на беспозвоночных; благодаря этому антигенеалогическому методу возник ряд недостатков, которые даже и до сих пор полностью не устранены. Так, при определении в сомнительных случаях зародышевых листков слишком часто прибегают к чисто топографическим признакам; говорят, например, об энтодерме у Orthonectidae и Dicyemidae, тогда как речь идет о клетках размножения, которые лежат под кожным слоем тела. Эд. ван Бенеден*** выставляет даже топографическое положение как единственный принцип определения зародышевых листков, как это следует
* В реферате в Журн. Мин. нар. просв., апрель 1869, стр.. 179. Неверно, когда Ковалевский (Studien uber Wurmer und Arthropoden, стр. 1), основываясь на этом моем высказывании, приписывает мне взгляд, как будто бы я не допускаю «никакого сравнения между зародышевыми листками позвоночных и беспозвоночных».
** Embryol. Studien an Wurmer u. Arthropoden. Mem. de 1'Acad. Sciences de St.-Petersbourg, т. XVI, № 12, 1871. Ковалевский пишет на стр. 5: «Зародышевые листки, которые я принимаю, имеют только то общее с описанными Мечниковым, что они зародышевые листки». Несправедливость этого резкого выражения вытекает из сравнения наших данных; в последнее время Ковалевский сам исследовал развитие скорпиона и убедился, что описанные мною зародышевые листки совершенно совпадают с его описанием. Если раньше нижний листок насекомых он не сравнивал с кишечно-слизистым листком позвоночных и, следовательно, также скорпиона, то эти взгляды им давно оставлены. Я должен приписать неправильному освещению моих данных со стороны Ковалевского то обстоятельство, что мое участие в вопросе о зародышевых листках обычно совершенно игнорируется.
*** См. его работы о Dicyemidae в Bull, de 1'Acad. Sciences de Belgique, 1876, и Archives de Biol., т. III, 1882, стр. 222, а также работы Жюлена, Гетте и других.
214
из следующих слов: «Nous appelons entoderme la couche ou la masse cellulaire enveloppee quels que soient, du reste, les tissus qui en derivent»*. Поэтому следует признать значительным шагом вперед, когда Геккель** впервые отчетливо сформулировал взгляд, согласно которому зародышевые листки, или по крайней мере два главных листка, следует рассматривать как первичные органы. Таким, путем был получен прочный и притом чисто генеалогический отправной пункт. С этой точки зрения то или иное образование лишь тогда могло быть определено как энтодерма, если оно совмещало несколько признаков этого первичного органа, а не только совпадало с ним по топографическому положению. Если бы, например, внутренняя клеточная масса Orthonectidae выполняла пищеварительные функции, то несомненно она была бы единодушно признана за энтодерму; но при существующих отношениях, когда она представляет собой исключительно половые клетки, это сравнение не может быть принято. Главное затруднение заключается в том, что при генеалогической трактовке проблемы зародышевых листков отсутствует прочный фундамент, так как нам неизвестно происхождение многоклеточных. Чтобы получить об этом представление, надо начать с гипотез, которые согласовались бы с наибольшим количеством фактического материала. Как эволюционное учение (Descendenzlehre) из чистой гипотезы превратилось в прочно обоснованную теорию после того как выяснилось, что при ее помощи может быть объяснена масса явлений и что никакие факты ей не противоречат, так же должны стать теорией и те гипотетические взгляды о происхождении первичных органов, которые полнее всего смогут быть приведены в соответствие с нашими положительными знаниями.
Хотя переходы между Protozoa и Metazoa среди современных животных, повидимому, больше не существуют, однако этот пробел в наших знаниях пытались в какой-то мере заполнить путем гипотетических построений. Можно допустить два пути, при помощи которых подобный переход мог бы осуществиться: или путем дифференцировки протоплазмы вокруг отдельных ядер у многоядерных простейших, или путем соединения многих особей колонии простейших в многоклеточное целое. В предыдущие десятилетия часто предполагали близкое родство между Ciliata и Turbellaria, а именно между их личинками, и поэтому казалось естественным между теми и другими принять наличие кровного родства и построить на этом гипотезу о происхождении Metazoa. Так это и произошло: многие исследователи высказались в пользу превращения многоядер-
* Мы называем энтодермой слой или покрытое [оболочкой] клеточное скопление, каковы бы ни были в конце концов происходящие из него ткани (фр.). — Ред.
** Studien zur Gastraea Theorie, 1877, стр. 258.
ных Opalina-подобных инфузорий в многоклеточных, и Иеринг*, а в последнее время Сэвиль Кент ** защищали этот взгляд. С этой точки зрения следует ротовое отверстие и порошицу инфузорий считать гомологичными одноименным органам многоклеточных; Иеринг даже думает, что воднососудистая система 119 последних может быть сведена к сократительной вакуоли простейших. Из всей совокупности процессов развития с этой гипотезой лучше всего согласуется образование бластодермы у насекомых. С. Кент действительно ссылается на первые эмбриональные стадии Aphidae. Эти выводы отчетливо доказывают, что подобная гипотеза не может считаться правильной. В то время как все процессы развития низших Metazoa ею игнорируются, она согласуется с образованием бластодермы у насекомых, т. е. у тех животных, которые во всяком случае вторично изменены. Но и среди последних имеются формы, которые противоречат гипотезе. Например, подуры, т. е. насекомые, которые занимают низшее положение во всем классе, в отношении процесса дробления яйца соответствуют многоножкам. Ввиду этих обстоятельств, гомология между ртом, порошицей и водной системой инфузорий и многоклеточных представляется совершенно необоснованной. С другой стороны, гипотеза перехода колонии жгутиконосцев в первичных Metazoa может объяснить большинство важнейших явлений; с этой точки зрения дробление яйца, к тому же примитивное, тотальное, может быть сведено к делению жгутиконосцев при образовании колонии, тогда как жгутиковый характер клеток многих бластул объясняется наследованием от жгутиконосцев. Если эта гипотеза препятствует сравнению рта и других «органов» простейших с одноименными частями Metazoa, то зато она помогает понять происхождение полового размножения, как это было показано впервые Бючли***. Понятно, что большинство эмбриологов, в том числе Р. Ланкэстер, Бальфур, высказались в пользу этой второй гипотезы, так что она, выдержав испытание, явилась базисом для дальнейших построений.
Естественно задать вопрос, нельзя ли несколько ближе представить себе особенности колонии жгутиковых, от которых произошли Metazoa. Бючли**** предполагает, что Metazoa происходят от двух корней: губок он выводит из колоний Choanoflagellata, а остальных многоклеточных — из колоний настоящих жгутиковых.
Не говоря уже о том, что существует слишком мало оснований для такого весьма рискованного утверждения, мы не можем
* Vergleichende Anatomie des Nervensystems und Phylogenie der Mollusken, 1877, стр. 21,
** A Manual of the Infusoria, т. II, 1881—1882, стр. 480. *** Studien uber die ersten Entwicklungsvorgange der Eizelle etc., 1876, стр. 219. **** «Bemerkungen zur Gastraea-Theorie». Morphol. Jahrd., 1883, стр. 124.
216
оставить без внимания то обстоятельство, что обе группы далеко не так резко обособлены друг от друга и что воротничок, образующий основное различие между ними, в некоторых случаях может совершенно исчезать. О родстве губок с кишечнополостными я еще будут говорить ниже. Независимо от того, имели или нет жгутиконосцы, давшие начало Metazoa, воротничок, во всяком случае несомненно, что они обладали способностью заглатывать твердые пищевые частицы, что доказывается распространением внутриклеточного пищеварения у низших Metazoa. Поэтому я не могу согласиться с Бючли, когда он форму питания для вопроса о происхождении Metazoa считает безразличной, «так как физиология питания среди Flagellata многократно меняется без связи с морфологией» (Bemerkungen z. Gastraea-Theorie, стр. 417). Я думаю скорее, что дальнейшая дифференцировка колонии жгутиконосцев, приводящая к образованию многоклеточных, ни в коем случае не явилась индифферентной в отношении типа питания. В то время как у некоторых выработались растительные пигменты, обеспечивающие ассимиляцию, другие Flagellata, сохранившие животное питание, достигли образования особых, специально для приема и переваривания пищи приспособленных индивидов. Что этот вывод; не является чисто дедуктивным, можно убедиться из сравнения таких колониальных жгутиконосцев, как Volvocineae и Protospongia.
Для дальнейших представлений было очень важно, если бы мы могли несколько более точно представить себе развитие гипотетических колоний жгутиконосцев. Что они должны были размножаться половым путем, вытекает уже из отношений, преобладающих у вольвоксовых. Как уже отмечалось выше, именно половое размножение относится к числу важных аргументов в пользу взгляда о происхождении Metazoa от жгутиконосцев. Что касается явлений деления, то мы знаем, что современные Flagellata в этом отношении ведут себя различно. У настоящих Flagellata господствует в общем продольное деление, хотя наряду с этим встречаются виды с поперечным делением, как, например, Phalansterium consociatum, по Ценковскому*, и Ph. digitatum, по Штейну**.
У Choanoflagellata, даже у близко родственных форм, тоже наблюдаются оба типа деления; так, согласно С. Кенту, Salpingoeca campanula размножается путем продольного деления ***, тогда как другие виды того же рода****, например S. inquillata, gracilis, — исключительно путем поперечного деления.
* Ueber Palmellaceen u. einige Flagellaten. Archiv f. mikroskop. Anat.,, т. VII, 1871, стр. 429, табл. XXIV, рис. 32.
** Der Organismus der Infusionsthiere, т. III, разд. 1, 1878, табл. VII, рис. 4,8.
*** A Manual of the Infusoria, т. III, 1880—1882, табл. IV, рис. 14. **** Там же, табл. VI, рис. 2—5, 27—29.
217
«Одновременное наличие продольного и поперечного деления у одной и той же формы до сих пор констатировано лишь у некоторых хламидомонад» *. Так как у колониальных животных способ деления особей играет важную роль для всей формы колонии, то важно узнать, как в этом отношении вели себя гипотетические Metazoo-Flagellata.
Как мы помним на основании выводов второй главы, три первых дробления совершаются в трех измерениях пространства и происходят одинаково у медуз с совершенно различными процессами развития (как у гипогенетических, так и. метагенетических, независимо от различий в образовании энтодермы и т. п.); так же протекают они у самых разных животных с полным дроблением. Поэтому представляется вполне достаточно обоснованным утверждение, что и у предков Metazoa имел место такой же тип деления. Это утверждение тем более вероятно, что к образованию зародыша может вести много путей, которые фактически и осуществляются как у растений, так и у животных с неравномерным дроблением. Даже образование бластулы может осуществляться без типичных трех первых дроблений. Мы видим, что у Volvox все эти дробления происходят в меридиональной плоскости, так что получается пластинчатый зародыш, напоминающий Gonium; при этом не образуется полости дробления и бластоцель формируется лишь путем последующего роста пластинки к одному полюсу **.
Если вообще использовать процесс дробления в генеалогическом смысле, то приведенные выше соображения представляются неопровержимыми. Они имеют аналогов в органическом мире, как показывают процессы деления у дробянок. В то время как большинство форм делится в поперечном направлении, бывают редкие исключения (например, найденный мною своеобразно ветвящийся и паразитирующий в Daphnia pulex вид Dendrobacterium ocellatum), у которых имеется лишь продольное деление.
Наряду с такими бактериями, у которых наблюдается лишь один единственный способ деления, имеются другие, где клетки делятся в двух меридиональных направлениях, как, например, микрококки гонорреи и, наконец, такие, как сарцины, где деления совершаются в трех измерениях пространства, т. е. сходны с полным дроблением большинства Metazoa, а также, как это предполагается, с гипотетическими Metazoo-Flagellata. Так как полость дробления в типичных случаях появляется уже после
* Butschli. Protozoa. Bronn's Klassen und Ordnungen, изд. 2, стр. 746.
** Сравнить сводку Бючли в Bronn's Klassen und Ordnungen. Protozoa, и его «Bemerkungen zur Gastraea-Theorie», стр. 423; далее Штейн «Organismus», т. III, 1, табл. XVII, XVIII. Я наблюдал первые стадии дробления у Volvox minor и проследил также образование пластинки с последующим превращением в открытый пузырь.
218
третьего дробления и зародыш очень рано превращается в бластулу, весьма вероятно, что и предки Metazoa плавали в виде бластулообразных колоний.
Если принять за основу для дальнейшего рассмотрения изложенные выше признаки Metazoo-Flagellata, то можно пролить, как мне кажется, некоторый свет на проблему происхождения первичных органов. Мы видели в третьей главе, что образование энтодермы у медуз происходит весьма различными способами. Если мы кратко вспомним эти способы, то должны будем констатировать, что энтодерма возникает или во многих точках зародыша, т. е. мультиполярно, или только в одной части последнего, т. е. концентрированно, или гипотропно. Мультиполярный способ возникновения энтодермы осуществляется: а) как мультиполярная иммиграция зародышевых клеток с поверхности внутрь эмбриона; b) как первичная деляминация путем поперечного деления бластодермальных клеток; с) как вторичная деляминация после предшествующего образования морулы или d) как смешанная деляминация, причем энтодермальные клетки образуются частью путем поперечного деления, частью путем миграции. Некоторые из этих способов образования нельзя резко разграничить, так как они связаны переходами. Гипотропное, или концентрированное, образование энтодермы осуществляется или путем иммиграции бластодермальных клеток нижнего конца личинки, или же путем настоящего впячивания, или гаструляции. К этому типу следует отнести и особую форму эпиболии, которая наблюдается лишь в виде вариации при неравномерном дроблении.
Спрашивается: с какой точки зрения можно объединить эти различные способы образования энтодермы и какая из существующих теорий происхождения первичных органов лучше всего объясняет фактически существующие отношения?
Легко видеть, что если принять гипотезу происхождения Metazoa от многоядерных Protozoa (инфузорий или солнечников и радиолярий), то остаются без всякого объяснения процессы, происходящие при эмбриональном развитии, как-то: иммиграция с поверхности, первичная деляминация, инвагинация. Эту гипотезу, как совершенно бесперспективную, следует поэтому отбросить, не входя в ее более подробную критику. Теория гастреи, как известно, разрешает очень многое путем сведения различных процессов развития к первичной инвагинации и часто помогает понять сложные процессы образования энтодермы, как, например, у позвоночных. Вместе с тем при объяснении деляминации она наталкивается на серьезные трудности, как это сразу признал Геккель еще при первой формулировке своей теории. «Наиболее тяжелое затруднение, — пишет он в своей монографии известковых губок (I, стр. 467, примечание), — заключается в том, что гаструла возникает из морулы двумя совершенно различными путями: в одних случаях
219
путем образования в моруле центральной полости прорыва образовавшейся таким путем пищеварительной полости наружу; в других случаях путем образования зародышевого пузыря, полого внутри шара, стенки которого состоят из одного слоя клеток, и впячивания этого зародышевого пузыря внутрь самого себя».
Геккель считал возможным преодолеть эти затруднения путем признания «вторичной фальсификации онтогенеза». В своей основной работе * он часто повторяет утверждение, что деляминация, если она действительно встречается в животном мире, представляет собой ценогенетический процесс, «который лишь вторично возник из палингенетического процесса инвагинации». Однако Геккель не входит ближе в рассмотрение возникновения подобной фальсификации. Это тем более вызывает недоумение, что сам Геккель чувствовал затруднение для своей теории в этом вопросе. Геккель и его школа (именно братья Гертвиги) долгое время вообще ставили под сомнение существование деляминации; в настоящее время таковая принимается, поскольку один из исследователей этой школы — О. Гаман** — сам наблюдал процесс деляминации у гидроидов (после того как он был описан многими предыдущими исследователями, как, например, Б. Оллмэном, Ф. Е. Шульце и мною). Он пытается преодолеть затруднение, непосредственно принимая деляминационную планулу за гаструлу и выводя ее путем ценогенеза из инвагинационной гаструлы. «Деляминация, — утверждает Гаман (стр. 504), — укороченная форма развития и во всяком случае выводится из инвагинации». «Ввиду повсеместного распространения гаструлы, — говорит он дальше, — остается в силе взгляд на планулу как на измененную гаструлу»; и еще: «... мы думаем, что имеем право говорить о гаструле и тогда, когда представляется вероятным, что отсутствие обоих образований (бластопора и пищеварительной полости) — вторичной природы. Поэтому мы называем планулой гаструлу, возникшую путем деляминации» ***. Эти утверждения высказываются без приведения доказательств, которые позволили бы понять, как инвагинация может, укорачиваясь, превратиться в деляминацию и что мог дать ценогенез для ее происхождения. Если между тем принять во внимание, что инвагинация концентрируется на одном конце зародыша и как раз у медуз ограничивается относительно небольшим участком бластодермы, тогда как первичная делями-
* Gastraea-Theorie, стр. 267, а также стр. 247, примечание; сравнить, также последнюю работу Геккеля: Ursprung u. Entwicklung d. thierischen Gewebe, 1884, стр. 50.
** Der Organismus der Hydropolypen. Jen. Zeitschr., т. XV, 1882. *** Геккель также называет различные личинки медуз, двуслойные планулы, личинки Geryonia и др. просто гаструлами, никак не обосновывая эта понимание и не приводя его в соответствие с его собственным определением гаструлы. См. его «System der Medusen», стр. 119, 298, 348 и т. д.
220
нация или мультиполярная иммиграция происходят в различных участках зародыша, то становится очевидным, что сведение первого способа образования на два последующих наталкивается на непреодолимые затруднения. Легко понять, как первоначально ограниченно распространенная инвагинация может постепенно распространяться на половину бластодермы, что часто и наблюдается у разных животных, или как однослойный зачаток энтодермы превращается в кучу связанных друг с другом клеток, которые затем обрастают эктодермой; но напротив — невозможно свести к укороченной инвагинации диффузное образование энтодермы, т. е. такое, когда энтодермальные клетки не соприкасаются с себе подобными, но чередуются с эктодермальными клетками (сравнить с развитием Aeginopsis) или же когда они возникают как центральные участки бластодермальных клеток.
Мультиполярную иммиграцию можно было бы еще попытаться свести к множественным впячиваниям, причем каждый первичный кишечник был бы представлен одной единственной клеткой! Но достаточно подобное предположение только сформулировать, чтобы увидеть его полнейшую несостоятельность. Но и таким путем мы не достигли бы многого, потому что первичная деляминация при этом осталась бы совершенно необъясненной. Если мы объяснение гаструляции у Craspedota хотели бы связать с эпиболическими явлениями, которые встречаются как вариации в развитии Polyxenia leucostyla, то нужно напомнить, что подобное образование тесно связано с неравномерностью бластомеров. Но когда у той же Polyxenia происходит равномерное дробление, то получаем вместо настоящей инвагинации (каковая у других Metazoa образуется из амфибластулы в результате равномерного дробления) скорее смешанную деляминацию, следовательно, такую форму развития, которая не может быть сведена к гаструляции. Этот пример показывает нам, что эпиболию у Polyxenia следует скорее рассматривать как изменение смешанной деляминации, которая в этом случае возникла вторично в связи с неравномерным дроблением. При таких обстоятельствах становится ненужным обсуждать подробнее отношения гаструляции ко вторичной деляминации, так как это не может привести ни к каким удовлетворительным результатам. Гораздо естественнее принять наличие связи между гаструляцией и гипотропной иммиграцией, как уже утверждали Клаус и другие; но мне представляется невозможным, без предварительного обсуждения ряда относящихся сюда вопросов, решить, какую форму следует рассматривать как исходную.
Хотя невозможность объяснения мультиполярного образования энтодермы следует рассматривать как сильнейшее возражение против теории гастреи, тем не менее ряд возражений против нее этим далеко еще не исчерпывается. Так как она была выдвинута в то время, когда предполагалось всеобщее распро-
221
странение полостного пищеварения и еще не было известно, что как раз низшие животные переваривают пищу внутриклеточно, то теперь она не соответствует больше нашим современным физиологическим представлениям 120. Согласно теории гастреи, между однослойной бластеей и двуслойной, снабженной «пищеварительной полостью» гастреей имеется целая пропасть, которая может быть легко заполнена, как только мы попробуем объяснить происхождение энтодермы с другой точки зрения. На дальнейших затруднениях, вытекающих из признания гомологии между всеми известными формами гаструл, нет необходимости здесь останавливаться, так как они непосредственно не связаны с вопросом о происхождении энтодермы и о них речь будет идти ниже.
Теория планулы Р. Ланкэстера* исходит непосредственно из наблюдавшихся при развитии Geryonia процессов и считает поперечное деление клеток бластодермы примитивным способом образования энтодермы. Если даже принять предположение Ланкэстера о сведении инвагинации к первичной деляминации, то остаются необъясненными такие случаи, когда, как при мультиполярной иммиграции, совсем не происходит деляминационного процесса, но энтодерма возникает из бластодермальных клеток, которые образуются продольным делением бластомеров и с различных точек поверхности проникают внутрь зародыша. Большое значение такого способа образования еще усиливается гипотропной иммиграцией, которая также основана на продольном делении бластодермальных клеток. Кроме того, против теории планулы, так же как и против теории гастреи, должно быть выдвинуто возражение, что обе эти теории стоят на точке зрения наличия полостного пищеварения у низших животных и, таким образом, находятся в противоречии с современным состоянием наших физиологических знаний. Р. Ланкэстер думает, что образование пищеварительной полости, в которую выделяется пищеварительный сок, произошло еще до развития энтодермы, что внутренние участки бластодермальных клеток функционировали как пищеварительные элементы, когда стенка была еще однослойной.
Все эти предположения оказываются совершенно неприемлемыми с тех пор, как стало известно, что внутриклеточное пищеварение сохраняется у многих низших организмов и даже обнаруживается у некоторых моллюсков (Phylliroё).
В то время как теории гастреи и планулы исходили из бластулы, состоящей из одинаковых клеток, Бальфур ** за основу своих собственных взглядов на переход от простейших к много-
* Notes on the Embryology and Classification of the Animal Kingdom. Quart. Journ. of Microscop. Science, N. S., т. XII, 1877, стр. 403.
** On the Morphology a. Syst. Posit. of Spongida. Quart. Journ. of Microscop. Science, N. S., т. XIX, стр. 103, и Handb. d. vergl. Embryologie, ubers. v. Vetter, т. I, 1880, стр. 143. '
222
клеточным кладет амфибластулу; поэтому его теорию можно просто назвать теорией амфибластулы. Так как ее следует рассматривать как модификацию теории гастреи, то остаются в силе все те возражения, которые направлены против последней. Прежде всего она оказывается не в состоянии объяснить такие формы развития, при которых энтодерма возникает не из одного зачатка, но из клеток, отделенных друг от друга лежащими в промежутках эктодермальными клетками (как, например, при мультиполярной иммиграции). Теория амфибластулы не дает также сколько-нибудь удовлетворительного объяснения первичной деляминации, происходящей путем поперечного деления бластомеров. Что касается приложимости теории Бальфура к губкам, то ее несостоятельность вытекает из наблюдавшихся еще прежними исследователями и мною фактов, показывающих, что энтодерма многих губок (именно известковых губок и Halisarcinae) представляет собой захватывающий пищу слой тела. Эти факты в последнее время подтвердил Гейдер* для Oscarella lobularis. Этим возражением устраняются и аргументы Бальфура, на которых он базирует утверждение об изолированном положении губок среди Metazoa.
К теории амфибластулы непосредственно примыкает теория плакулы Бючли **, и не только потому, что автор также высказывается в пользу изолированного происхождения губок, но прежде всего потому, что плакула во многих отношениях может рассматриваться как расплющенная амфибластула. Бючли признает недостатки других теорий в вопросе о генеалогии зародышевых листков и делает попытку чисто схематически объединить инвагинацию и деляминацию под общим углом зрения, причем он выводит оба способа образования энтодермы из изменений первичной плакулы. Отказываясь от шарообразной колонии простейших — исходного пункта прежних построений, Бючли принимает в качестве исходной формы однослойную Gonium-подобную пластинку, которую я в дальнейшем ради удобства буду называть проплакулой. «Мне представляется приемлемым, — пишет Бючли (стр. 419), — что двуслойное состояние сначала возникло в колонии простейших, клетки которой располагались в одной плоскости рядом друг с другом, образуя однослойную пластинку. Если все клетки разделятся параллельно поверхности пластинки, то возникнет двуслойная пластинка, оба слоя которой, вероятно, еще сначала не будут обнаруживать дифференцировки. Для простоты понимания мы будем эту стадию двуслойной пластинки называть плакулой». В то время как подобная плакула, прини-
* Zur Metamorphose von Oscarella lobularis. Arbeiten d. zool. Inst. d. Univ. zu Wien, т. VI, вып. 2, 1886, стр. 227. В этой важной работе Гейдер придает слишком большое значение гаструляции Oscarella и упускает в своих теоретических рассуждениях способы образования энтодермы у других губок.
** Bemerkungen zur Gastraea-Theorie, Morphol. Jahrb., т. IX, стр. 415.
223
мая трубковидную форму, превращается в гаструлу, из проплакулы в результате вторично запоздавшего клеточного деления возникает деляминационная бластула. Постулатом для подобного предположения является принципиальное различие форм бластул, которые должны рассматриваться или как вздувшиеся плакулы, или как ставшие шаровидными проплакулы. В истории развития современных животных плакула, согласна Бючли, представлена уплощенными бластулами Cucullanus (по Бючли), Rhabdonema (по Гетте), Lumbricus, Chiton (по Ковалевскому), Phoronis, Ascidia mentula, среди взрослых животных — у Trichoplax adhaerens F. E. Sch. Бючли, однако, не замечает, что уплощенные бластулы указанных выше Metazoa, поскольку известно их развитие, лишь по внешней форме, а не существенными морфологическими чертами отвечают его плакуле, так как у них двуслойность отнюдь не достигается путем «параллельного плоскости пластинки» деления, которое для плакулы является основным. У Phoronis, Ascidia, принципиально, конечно, и у других из указанных выше животных плакулообразная стадия возникает путем сплющивания первоначальной более или менее пузыревидной бластулы, а не наоборот, как этого требует теория. Последняя сравнивает деляминацию Geryonidae, осуществляющуюся путем поперечного деления бластомеров, с плакулами и амфибластулами других животных; у последних, следовательно, также должно происходить поперечное деление бластомеров, что фактически, совершенно определенно, не имеет места. Энтодермальные клетки пластинчатых бластул не возникают непосредственно из лежащих над ними эктодермальных клеток, но из клеток, лежащих рядом с ними. Мы приходим к заключению, что стадия плакулы в истории развития современных животных с равномерным дроблением совершенно не встречается. Еще с меньшим правом можно принимать за таковые некоторые стадии эмбрионального развития Ctenophora, где над пластинчатым зачатком энтодермы лежит прерывающийся эктодермальный слой*; в этом вряд ли кто-либо признает первичную стадию, которую можно было бы положить в основу морфологического обобщения.
Если, возвращаясь к процессам развития, в поперечном делении четырех первых бластомеров по пути к восьмиклеточной стадии, видеть преходящую стадию плакулы, то можно возразить, что этим ничего не объясняется, так как, во-первых, восьмиклеточная стадия у деляминирующих Geryonidae совершенно сходна с той, которая имеется у Acraspeda с их инвагинацией, и, во-вторых, деляминация путем поперечного деления не могла бы быть отнесена к стадии плакулы, поскольку эта стадия уже осуществилась раньше.
* Сравнить с моими «Vergleichend-embryologische Studien, 4» в Zeitschr. f. wiss. Zool., т. XLII, 1885, стр. 650.
224
Даже если допустить, что теория плакулы удовлетворительно объясняет образование инвагинационной гаструлы в результате поперечного деления проплакулы, то остаются незатронутыми такие явления, как образование энтодермы путем мультиполярной или локальной иммиграции с предшествующим продольным делением клеток зародыша.
Согласно теории Бючли, у животных с деляминацией должен иметь место такой же способ формирования бластулы, как у Volvox, т. е. образование бластулы с предшествующей стадией проплакулы (напоминающей Gonium). Бючли считает возможным действительно принять наличие таковой у Geryonia probos-cidalis согласно данным Фоля (стр. 423 цит. выше работы). На самом деле шестнадцатиклеточный зародыш Geryonidae уже представляет собой готовую типичную бластулу, возникшую из восьмиклеточной стадии таким же путем, как и у медуз с другими способами образования энтодермы; восьмиклеточный зародыш в свою очередь образуется путем такого же экваториального дробления, как и у других медуз и вообще у большинства Metazoa с равномерным дроблением. Среди последних наибольшее сходство с вольвоксом (в отношении предшествующих бластуле стадий) обнаруживает, по Ф. Е. Шульце *, Sycandra raphanus, так как у этой губки имеется восьмиклеточная пластинчатая стадия; однако это ни в какой мере не может служить в пользу теории плакулы, ибо как раз Sycandra характеризуется инвагинационной гаструлой.
Вопрос о морфологии интересного Trichoplax adhaerens F. Е. Sch. ** и его отношение к плакуле в настоящее время еще нельзя обсуждать серьезно. Нельзя даже приближенно определить, какое значение имеют различные слои тканей названного животного; на основании гистологических различий обеих поверхностей тела можно столь же мало делать заключение о различии зародышевых листков, как у губок, где тот же зародышевый листок — энтодерма — выступает то в форме жгутикового эпителия камер, то как совершенно уплощенный эпителий собирательных полостей, например у Oscarella lobularis (по Гейдеру). Затруднения увеличиваются не только вследствие незнания процессов развития, но и вследствие того, что не существует никаких данных для определения физиологического значения слоев тела Trichoplax. Благодаря любезности профессоров Ф. Е. Шульца и Клауса, я изучал Trichoplax в 1883 г. в Граце и Вене, причем мог полностью подтвердить гистологические данные первого из двух названных выше исследователей; мои опыты питания Trichoplax дали отрицательный результат, поскольку животные вообще не заглатывали пищи, так что создается впечатление, что Trichoplax приспособился к жидкой пище121.
226
* Die Metamorphose von Sycandra raphanus. Zeitschr. f. wiss. Zool., т. XXI, 1878.
** Trichoplax adhaerens. Zool. Anz., VI Jahrg., 1883, стр. 92
Бючли думает, что теория плакулы и в физиологическом отношении более приемлема, чем другие критикуемые им взгляды. Выдвигаемые им против этих теорий возражения кажутся мне безусловно оправданными. «Мне представляется очень существенным, — пишет Бючли (указ, соч., стр. 416), — что изменения принимаемых нами форм должны быть постепенными, не скачкообразными, а также действительно полезными». «Особенно в последнем отношении, я думаю, — добавляет Бючли, — что развиваемый новый взгляд имеет некоторые преимущества перед прежними». Когда он подходит к объяснению происхождения плакулы физиологически, то достаточных оснований не приводит. «К сожалению, я не в состоянии, — утверждает сам Бючли (стр. 419, примечание), — привести правдоподобное объяснение полезности для происхождения двуслойности пластинки».
Если мы просмотрим вновь все сказанное выше об обсуждавшихся теориях, то увидим, что они не в состоянии объединить под общим углом зрения всю сумму известного эмбриологического материала и что они, кроме того, страдают отсутствием физиологических объяснений. Поэтому должен быть найден другой путь. По ходу моих исследований над губками * я очень осторожно сделал некоторые замечания, которые, по-моему, согласуются с полученными данными об образовании энтодермы у низших Metazoa, а также соответствуют ставшим известными явлениям внутриклеточного пищеварения. Я думал, что энтодерма не возникла сразу в форме трубчатого желудка с терминальным отверстием, как мы это находим у гаструлы, но что эти образования имеют за собой длительный процесс исторического развития, который проявляется в формировании сплошной переваривающей внутриклеточно паренхимы. Последняя образовалась тоже не сразу, а постепенно, причем поверхностные бластодермальные клетки мигрировали внутрь бластоцеля.
В конце концов возникла двуслойная паренхимелла, которая путем укорочения эмбрионального процесса и прогрессирующей дифференцировки пищеварительного аппарата превратилась в гаструлу. При невозможности в то время (в 1879 г.) сослаться на каких-либо высоко дифференцированных жгутиконосцев с животным способом питания я пытался найти некоторые точки опоры в размножении Volvox и писал следующее: «По-моему, своевременны поиски таких низших существ, у которых питательные клетки после заглатывания пищи покидают свое обычное место, чтобы с поверхности «колонии» проникнуть во внутреннюю полость» (указ, соч, стр. 382). Вскоре после
* Spongiologische Studien. Zeitschr. f. wiss. Zool., т. XXXII, 1879, стр. 374. Эти же взгляды я высказал в 1877 г. в трех статьях на русском языке в. Изв. Новоросс. О-ва естествоисп., т. V.
226
этого (июль 1880 г.) Сэвиль Кент * открыл чрезвычайно интересную колониальную форму Flagellata, которую он ввел в науку под именем Protospongia Hackelii. Среди первоначально правильно и периферически расположенных особей колонии некоторые принимали выраженную амебоидную форму (см. гравированный рис. 1,а)122 и внедрялись внутрь студенистой массы, связывающей членов колонии. Здесь амебоидные особи и оставались, приступая к делениям и другим изменениям, которые С. Кент принимает за споруляцию (см. гравированный рис. 2). Находятся ли переход в амебоидную форму и внедрение в студенистое вещество в какой-либо связи с питанием, в настоящее время утверждать нельзя, так как эти отношения еще совер-
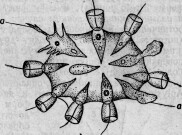
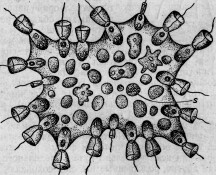
шенно не выяснены. Ввиду того факта, что и споруляция представляет еще много неясного, я хотел бы высказать предположение, не являются ли многочисленные наблюдавшиеся Кентом зер-
* A Manual of the Infusoria, т. I, 1880—1881, стр. 363, Атлас, табл. X, рис. 20—30.
227
Рис. 2.
Рис. 1.
нышки или запасными питательными веществами, или же непосредственно заглоченными частичками пищи. Было бы исключительно интересно ближе исследовать род Protospongia (так же как и вторую описанную Оксли, состоящую из многих особей форму Protospongia), имея в виду именно процессы питания и размножения. Во всяком случае можно принять, что этот воротничковый жгутиконосец обладает двумя формами особей (которые естественно переходят друг в друга): жгутиковыми и амебоидными; последние могут внедряться с различных точек поверхности в общую студенистую массу. Нельзя отказать в большом сходстве Protospongia с некоторыми двуслойными личинками губок, например, с описанной Ф. Е. Шульце личинкой Aplysina stilphurea *, так что в дифференцировке амебоидных
форм особей мы можем предположительно видеть первый шаг к образованию энтодермы 123.
Во всяком случае я думаю, что особенности Protospongia легче согласовать с моими взглядами (которые некоторыми авторами называются теорией паренхимеллы), чем со всеми выше рассмотренными теориями других исследователей. Как относится, однако, теория паренхимеллы к фактам из области истории развития вообще и к изложенной выше эмбриологии медуз в особенности? Чтобы осветить этот вопрос, следует сначала вспомнить, что я a priori утверждал в отношении явлений размножения гипотетических Metazoo-Flagellata.
Из всеобщего распространения трех первых делений при дроблении, которые совершаются в трех измерениях пространства (сагиттально, фронтально, экваториально) и, следовательно, представляют собой поперечные и продольные деления, я сделал заключение, что и предки Metazoa делились в этих же различных направлениях. Постепенно направления деления закрепились таким образом, что одни формы размножались преимущественно или исключительно путем продольного деления, в то время как другие, родственные им, еще сохраняли поперечное деление. Что сходные явления действительно могут иметь место, доказывают приведенные выше виды Salpingoeca. Итак, мы должны принять, что у наших гипотетических Metazoo-Flagellata некоторые поверхностные клетки становились амебоидными и внедрялись внутрь колонии, совершенно так же, как сегодня мы это наблюдаем у Protospongia; что наряду с этим другие особи
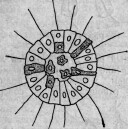
Рис. 3.
* Zeitschr. f. wiss. Zool. т. XXX, 1878, стр. 416, табл. XXIV, рис. 30.
228
делились в поперечном направлении, причем центральный продукт деления попадал внутрь, тогда как периферический сохранял свое поверхностное положение. Гравированный рис. 3 представляет эти отношения в схематическом виде. (Возможно, что наряду с поперечным и продольным делениями встречались и такие
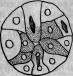

Рис. 4.
Рис. 5.
Рис. 6.
формы, которые делились в косом направлении, как это и в настоящее время происходит у некоторых простейших.)
Эмбриологически этот двойной способ образования энтодермы путем внедрения целых клеток и отшнуровывания цент-тральных продуктов деления осуществляется в тех случаях, когда, как, например, у Polyxenia leucostyla, имеется смешанная деляминация (гравированный рис. 4). В то время как у некоторых форм выступает на первый план поперечное деление (гравированный рис. 5), у других господствует продольное деление
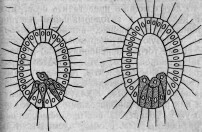
Рис. 7.
Рис. 8.
Рис. 9.
и, следовательно, иммиграция целых поверхностных клеток (гравированный рис. 6). Так произошли от смешанной деляминации, с одной стороны, первичная деляминация, а с другой — мультиполярная иммиграция. Вторичную деляминацию следует рассматривать лишь как небольшую модификацию смешанной, так как она отличается только несколько более поздним наступлением различий между обеими группами клеток.
229
Следует несомненно принять, что мультиполярное образование энтодермы является более первичной формой, чем концентрированное (гравированный рис. 7), так как противоположное допущение наталкивается на большие затруднения, как мы это уже видели выше. Переход от мультиполярной к гипотропной иммиграции, напротив, легко себе представить, тем более что последняя встречается преимущественно у сильно подвижных бластул, тогда как первая имеет место у неподвижных или малоподвижных эмбрионов. При обсуждении вопроса об образовании энтодермы у метагенетических медуз я подчеркивал, что продольное деление представляет собою правило, поперечное же деление встречается как очень редкое исключение.
Если концентрированная иммиграция может быть без труда выведена из мультиполярной, то, с другой стороны, нетрудно инвагинацию свести к первой из них. Мы видели в третьей главе, что образование энтодермы у Laodice cruciata отличается от того же процесса у других метагенетических Craspedota тем, что на заднем полюсе бластулы выступает ряд прозрачных клеток (табл. IV, рис. 24—28) *, которые не все сразу, но постепенно вдвигаются в бластоцель (гравированный рис. 8). После этого образуется паренхиматозная энтодерма, в которой возникает полость, причем и сама энтодерма приобретает характер эпителия. Исходная стадия с прозрачными клетками обнаруживает чрезвычайное сходство с бластулой Nausithoё marginata, которая также характеризуется дифференцировкой гипотропного ряда клеток (табл. X, рис. 12) *. Если мы допустим дальнейшее укорочение хода развития, то легко можно представить себе, что дифференцированные на поверхности энтодермальные клетки, вместо того чтобы мигрировать поодиночке, впячиваются совместно и прямым путем образуют энтодермальную полость (гравированный рис. 9). После того как первоначально дифференцированные клетки инвагинировались, этот процесс переходит на соседние бластодермальные элементы, благодаря чему впячивающийся мешок понемногу увеличивается. Дальнейший мыслимый процесс сокращения развития у медуз не осуществился; он представляется как ранняя дифференцировка энтодермальных клеток, так что уже у бластулы все предназначенные к инвагинации клетки различимы как особые элементы. Как следствие такого укорочения следует рассматривать, наряду с другими, и пластинчатые бластулы с двумя более или менее одинаковыми зародышевыми слоями (как, например, у Lumbricus, асцидий и т. д.). Очевидно, что дальнейшие шаги в ранней дифференцировке энтодермы приведут к амфибластуле; это будет замечено уже во время дробления и, наконец, может быть выражено даже в структуре яйца.
* Эти рисунки, относящиеся к тексту других глав «Эмбриологических исследований над медузами», нами не воспроизводятся. См. их в «Избранных биологических произведениях», М., 1950. — Ред.
230
Если амфигаструла может возникнуть из архигаструлы в (результате ранней эмбриональной дифференцировки, как это общепринято со времени Ковалевского (Euaxes), то, с другой стороны, несомненно, что сходная форма может образоваться и путем неравномерного дробления и смешанной деляминации, как мы видели на примере Polyxenia leucostyla. Приведенный случай учит нас, что амфигаструлы могут возникать двумя разными путями из двух разных исходных стадий и доказывает, следовательно, полифилию указанной эмбриональной формы.
С изложенной точки зрения энтодерма должна рассматриваться как сумма клеток, которые первоначально образовались из бластулы путем иммиграции или отделения и сгруппировались в различно развитое скопление амебоидных элементов.
Гастральная полость, так же как и ротовое отверстие, должны рассматриваться как позднейшие приобретения, которые закрепились с помощью укорачивания эмбрионального развития и привели к дефинитивному образованию гаструлы.
Бальфур*, который высказался против теории паранхимеллы, признает, однако, сам, что она «хорошо согласуется с онтогенезом низших Hydrozoa». Теперь, когда стало известным большое количество фактов, это соответствие выступает еще отчетливее. Когда, однако, Бальфур говорит, что устанавливаемый теорией паренхимеллы переход от Protozoa к многоклеточному состоянию ему «представляется невероятным», то я ничего не могу возразить против его критики, так как свое утверждение он ничем не подкрепляет. Гораздо более обстоятельны возражения Бючли, которые относятся исключительно к физиологической стороне вопроса. После краткого изложения моих взглядов он добавляет следующие критические замечания (указ, соч., стр. 418): «Мне кажется, что иммиграция энтодермальных клеток, функцией которых является захват пищи, не может рассматриваться как преимущество. Без одновременного образования рта, которое в гипотезе Ланкэстера не мотивировано и непонятно, иммиграция энтодермальных клеток, по-моему, не была бы полезна, потому что они сами, так сказать, создавали бы перед собою препятствие».
Когда я опубликовал мою иммиграционную теорию образования энтодермы **, я уже ссылался на вольвокс, у которого размножающиеся особи направляются с поверхности колонии в ее глубину.
Теперь я могу еще в качестве доказательства привлечь внедрение особей у Protospongia, которое, конечно, представляется полезным для жизни колонии в целом. Кроме того, можно привести еще априорные соображения. Между особями колонии должно было сначала возникнуть неравенство, которое привело
* Handb. d. vergl. Embryol., т. II, стр. 308. ** Zeitschr. f. wiss. Zool., т. XXXII, стр. 382.
231
к дальнейшей дифференцировке: в то время как некоторые особи преимущественно заглатывали пищу, другие были более приспособлены к движению и привлечению пищи. Для подвижных колоний должно было представлять определенную пользу то обстоятельство, что снабженные пищевыми частицами, перегруженные и поэтому более тяжелые особи не оставались на периферии, но перемещались возможно ближе к центру. Другое преимущество заключалось еще в том, что питающиеся особи были поставлены в условия, позволяющие возможно совершеннее выполнить свою функцию. Известно, что многие Flagellata переходят из состояния монады в амебоидное, в котором они как раз и принимают пищу. Я позволю себе сослаться на изложение Бючли *. «У многих Monadina находим локализацию приема пищи на определенном ротовом участке, который почти всегда располагается у основания жгутика. Если в этом факте... нельзя сомневаться, то, с другой стороны, нельзя и отрицать, что некоторые относящиеся сюда формы, у которых наблюдается иногда переход в саркодообразное состояние, в это время могут принимать пищу и другими участками тела». Далее легко представить, что питавшиеся особи чаще размножались, так что могла образоваться известная зависимость между более интенсивным питанием и размножением. Еще одно основание для наличия иммиграции заключается в невозможности поверхностного расширения колонии свыше известной границы, что именно у плавающих колоний должно иметь большое значение. Так как увеличение числа особей делало возможным повышенную активность, то было несомненно полезно, когда клетки, не находившие больше места на поверхности, приспособились к существованию внутри колонии. Долгое время особи одной колонии отличались друг от друга, вероятно, лишь количественно: локомоторные клетки при помощи движения жгутиков добывали пищевые частицы и наиболее мелкие из них частично заглатывали сами. И в настоящее время у некоторых Coelenterata можно наблюдать случайное заглатывание пищи эктодермальными клетками**. Внутренние амебоидные особи были, напротив, способны заглатывать и более крупные частицы, которые не могли быть захвачены локомоторными элементами. Вероятно, амебоидные особи при этом почти достигали периферии и через многочисленные поры поверхностного слоя захватывали частицы, соприкасавшиеся с поверхностью колонии.
В качестве аналогичного явления можно привести захватывание посторонних частиц амебоидными клетками перезимовывающих Halisarca pontica, которые теряют жгутиковые каналы, а также заглатывание мезодермальными клетками у многих
* Protozoa, в Bronn's Klassen und Ordnungen, 2-е изд., стр. 695. ** Untersuchungen uber die intracellulare Verdauung, в Arbeiten des zool. Inst. d. Univ. zu Wien, т. V, 1883, стр. 142.
232
низших животных. Постепенно дифференцировка в намеченном направлении достигала все большей степени; локомоторные клетки более или менее теряли функцию захвата пищи, которая концентрировалась в амебоидных фагоцитах; случайные тончайшие поры между локомоторными элементами могли увеличиться и превратиться в отверстия, которые в таком большом количестве встречаются на поверхности губок. По мере нарастающей индивидуализации колонии (индивидуумы второго порядка), поверхностные особи дифференцировались в эктодерму, или кинобласт, тогда как амебоидные внутренние особи — в фагоцитобласт (паренхима или мезо-энтодерма). Клетки последней, в случае невозможности овладеть более крупными пищевыми частицами, образовывали вокруг них плазмодии, подобно тому как это происходит в настоящее время в энтодерме сифонофор или в мезодерме многих животных. Повышенная активность снабженного двумя первичными органами многоклеточного приводила к удовлетворению потребности в пище; при этом, вероятно, поедались и более крупные растительные и животные организмы. Чтобы сделать это возможным, должны были возникнуть одно или несколько входных отверстий, которые могли привести к образованию рта.
Для описанной выше дифференцировки однородных особей на локомоторных и фагоцитов я могу привести еще следующие аналогии: у низших Coelenterata, как, например, у гидроидных полипов, некоторых медуз (например, Oceania, Phialidium, Cunina) вся энтодермальная стенка гастроваскулярной системы одинаково способна поглощать питательные частицы путем внутриклеточного пищеварения; все энтодермальные клетки являются фагоцитами, а также локомоторными элементами в тех случаях, когда они снабжены жгутиками. У других Coelenterata, как, например, у Aglaura среди Craspedota, у Siphonophora и Ctenophora, энтодерма, напротив, дифференцируется в амебоидные эпителиальные клетки, которые только одни поглощают пищу, и жгутиковые эпителиальные клетки, служащие для поддержания тока воды, но не способные заглатывать посторонние частицы. У губок мы видим совершенно сходное явление: у многих представителей жгутиковые клетки энтодермы действуют, как фагоциты, в то время как у некоторых кремневых губок эта роль переходит исключительно к амебоидным клеткам, так как энтодермальные жгутиковые клетки служат лишь для движения тока воды.
Что иммиграция особей с поверхности колонии во внутреннее пространство (которое, вероятно, было выполнено студенистым веществом) не исключает возможности поперечного деления других особей, понятно само собой, так же как и то обстоятельство, что при наступившей дифференцировке периферический продукт деления мог оставаться на месте, тогда как центрально расположенная дочерняя особь оказывалась внутри колонии в качестве
233
фагоцита. Переходную форму между жгутиконосцами и многоклеточными, от которой произошли последние, я назвал прежде паренхимеллой. Теперь я хотел бы переименовать ее в фагоцителлу (Phagocytella), так как это название указывает на очень характерную особенность этой формы. Пока можно предположить, что фагоцителла обладала двумя первичными органами, кинобластом и фагоцитобластом, которые, однако, еще не так резко были отделены друг от друга, как зародышевые листки большинства Metazoa; вероятно, еще долгое время продолжалось пополнение фагоцитобласта внедряющимися клетками кинобласта. В отношении истории развития можно сказать, что яйца фагоцителлы (она должна была обладать половым размножением) претерпевали равномерное дробление, причем бластомеры делились в трех измерениях пространства, и что рано возникал бластоцель, который затем постепенно заполнялся отдельными мигрирующими целыми клетками и центральными продуктами деления клеток.
Теория фагоцителлы согласуется с нашими знаниями в области морфологии и физиологии губок. Эмбриология различных губок обнаруживает разнообразие, почти превосходящее таковое у медуз; хотя они до сих пор еще далеко не достаточно изучены, но уже и сейчас можно утверждать, что в ходе эмбрионального развития этого типа животных встречаются как процессы иммиграции, так и деляминация и гаструляция.
В целом сказанное выше об эмбриологии медуз приложимо также и к разным способам образования энтодермы у губок. Первичные органы последних сохранились в относительно примитивном состоянии, так что распространение схемы зародышевых листков на слои тела губок представляет большие трудности, как я это неоднократно испытывал сам. Так как «энтодерма» часто переходит в «мезодерму», так что оба образования находятся в самой теснейшей связи *, я думаю, что оба слоя вместе следует рассматривать как фагоцитобласт. Что образующие фагоцитобласт клетки не возникают все сразу, но часто путем последующей миграции добавляются из эктодермы (например, у Halisarca), ни в какой мере не опорочивает нашего взгляда. Организация губок не представляет ничего такого, на чем вместе с Бальфуром и Бючли можно было бы обосновать их отделение от прочих Metazoa, так что я в этом отношении целиком могу стать на сторону большинства спонгиологов и в том числе К. Гейдера **.
Отсутствие одного ротового отверстия или же наличие многих входных пор можно считать весьма значительными различиями только в том случае, если этим образованиям приписывать неправильное генеалогическое значение; с точки зрения теории фагоцителлы указанные особенности тела губок, напротив, легко
* Zeitschr. f. wiss. Zool., т. XXXII, стр. 374. ** Oscarella lobularis. См. указ, на стр. 435 соч., стр. 56. 234
объяснимы. Преобладающая роль амебоидных клеток при приеме пищи была бы лишь в том случае странной, если мы у губок могли бы говорить о дифференцированной мезодерме; но так как губки обладают еще фагоцитобластом, который должен рассматриваться как общий зачаток энто- и мезодермы, то указанный факт окажется, напротив, весьма понятным.
После того как я опубликовал мои взгляды в общих чертах в «Spongiologische Studien» *, Бальфур пришел уже в следующем году к выводам о развитии кишечнополостных, которые согласовались с основными положениями теории фагоцителлы. «Как это ни может показаться парадоксальным, — пишет английский эмбриолог **, — я не считаю невозможным, что кишечнополостные на самом деле имели предка, у которого пищеварительный канал был представлен физиологически сплошной массой амебоидных клеток. Этот предок был, может быть, кроме того, общим и для турбеллярий».
Представляется удивительным, что при подобном взгляде Бальфур столь решительно склоняется на сторону теории гастреи, а теорию паренхимеллы считает для себя неприемлемой. В последнее время Гетте*** повторил взгляд о близком родстве исходной формы кишечнополостных и турбеллярий (Acoela), так же как и о паренхиматозном характере энтодермы у первичных Metazoa, не ссылаясь, впрочем, ни на Бальфура, ни на меня. Во всяком случае, из этого можно видеть, что число исследователей, чувствующих себя вынужденными принимать наличие фагоцителла-подобного этапа, постепенно увеличивается. Хотя новейшие исследования по анатомии Acoela (Клейненберга, Переяславцевой, И. Делажа и мои собственные) исправляют данные Граффа об отсутствии у этих турбеллярий мезодермы и нервной системы, тем не менее остается в силе тот факт, что органы пищеварения Acoela сохраняют очень примитивный характер. Если в последнее время утверждали, что названные черви обладают выраженной пищеварительной полостью, то я должен против этого возразить, что настоящие Acoela обнаруживают энтодермальный плазмодий, содержащий различной величины вакуоли, которые могут симулировать пищеварительную по-
* Уже в моей работе об органах пищеварения пресноводных турбеллярий (в «Изв. Новоросс. О-ва естествоисп.», т. V, 1877) я писал следующее: «Если в самом деле турбелларии по существу своему настоящие паренхиматики, то оказывается, что в этом отношении существует основное сходство между низшими представителями двух исходных типов многоклеточных животных (Coelenterata и Vermes), т. е. между губками и турбеллариями. Сравнивая личинки первых (в особенности описанные О. Шмидтом личинки Amorphina) с низшими представителями последних, можно придти к предположению, что оба названные класса скреплены между собою несравненно более тесным родством, чем это предполагалось прежде»124. ** Handb. d. vergl. Embryol.. т. I, 1880, стр. 172.
*** Abhandlungen z. Entwicklungsgeschichte d. Thiere, т. II, 1884, стр. 184 и сл.
235
лость. Эти отношения я исследовал у прозрачной пелагической формы из Мессины (где она также многократно изучалась Клейненбергом) и совершенно убедился в правильности этих данных. Из эмбриологических исследований г-жи Переяславцевой* и Репяхова** как будто вытекает, что после дробления у изучавшихся ими Acoela наступает стадия гаструлы; последний из названных авторов*** делает из этого вывод, что Acoela представляют собой регрессивно видоизмененных червей. Я хотел бы возразить, что образование гаструлы далеко не обязательно следует рассматривать как генеалогически первичный процесс; мы видели в эмбриологии медуз, как гаструлы возникают полифилетически из разных первичных образований (в одном случае как архигаструла Nausithoё и Pelagia, в другом — как эпиболическая гаструла у Polyxenia leucostyla), так что ее возникновение в процессе филогенеза легко можно представить себе как вторичное эмбриональное приспособление. Кроме того, я должен заметить, что окончательное суждение о гаструле Acoela сможет быть высказано лишь после опубликования полностью работ г-жи Переяславцевой и Репяхова 125.
Так как, согласно развиваемому мною взгляду, гаструлы могут возникать независимо в ходе эмбрионального развития, то благодаря этому преодолеваются и даже совершенно обходятся многие затруднения теории гастреи. В этой связи я отсылаю к моим «Сравнительно-эмбриологическим исследованиям»**** где я посильно эту сторону вопроса осветил. В последние годы Блохман***** и Седжвик****** пытались спасти теорию гастреи, приняв положение о том, что рот и анус — оба образуются из щелевидного бластопора. Они ссылаются при этом на исследования Бальфура над Peripatus и на данные, полученные у Aplysia и других Gasteropoda, и думают, что щелевидный бластопор, образующий рот и анус, принципиально может быть принят и для всех Metazoa. Главное затруднение теории гастреи таким путем, однако, не разрешается. С изложенной Седжвиком точки зрения, радиально построенные гаструлы иглокожих Pilidium и Polygordius должны рассматриваться как вторично сильно измененные личиночные формы. Эмбрионы же Gasteropoda, Peripatus, насекомых и червей с щелевидным бластопором с этой точки зрения представляют собой первичные состояния гаструлы. По этой же причине правильные бластулы
* Sur le developpement des Turbellaries. Zool. Anz., т. VIII, 1885, № 194, стр. 269.
** Там же, стр. 272.
*** В его диссертации на русском языке о Dinophilus, Одесса, 1886, стр. 47.
**** Zeitschr. f. wiss. Zool., 1882, стр. 286.
***** Beitrage z. Kenntn. d. Entwicklung d. Gasteropoden. Zeitschr. f. wiss. Zool., т. XXXVIII, 1883, стр. 406.
****** On the Origin of metameric Segmentation etc. Studies from the Morphol. Laboratory in the Univ. of Cambridge, т. II, 1884, ч. I, стр. 82.
236
названных выше первыми животных следует рассматривать как измененных потомков богатых желтком амфибластул других форм.
В генеалогии ануса, которая в цитированных выше теориях не получает достаточного объяснения, мы можем обнаружить те же ступени, которые приняли при образовании рта. У низших Metazoa мы видим возникновение двух (Ctenophora) или большего количества отверстий, служащих для выведения продуктов выделения, аналогично тому как у губок имеются многочисленные входные отверстия. У некоторых медуз Lafoeidae (Aequorea, Tima) имеются на особых папиллах многочисленные выделительные отверстия гастроваскулярной системы; некоторые Polyclada обладают сходными выделительными отверстиями на различных участках тела 126. У Cycloporus Ланг * наблюдал выталкивание через такие отверстия пищеварительного аппарата капелек жидкости с различно окрашенными включениями. Эта находка приобретает тем большее значение, что Polyclada обладают, кроме того, еще особой выделительной системой.
В то время как часть фагоцитобласта превратилась в энтодерму, в которой амебоидные клетки постепенно приобрели эпителиальный характер, другая часть этого первичного органа дала мезодерму.
Сначала последняя выступала в виде единичных подвижных клеток, которые действовали так же, как раньше фагоциты. Подобное отношение мы находим у многих кишечнополостных (у которых в некоторых случаях еще добавляются бродячие клетки из эктодермы, как, например, у кораллов, по Ковалевскому и Мариону), у иглокожих, многих червей и т. д. Таким образом, обнаруживается известная гомодинамия в развитии мезодермы, так как она может образовываться в различных случаях путем миграции клеток. Наряду с тем, что вся мезодерма или часть ее постоянно сохраняет первичное состояние фагоцитобласта, у разных животных образуются еще особые мезодермальные мешки, которые отделяются от энтодермы, причем их клеточные стенки в той или иной степени теряют фагоцитарную функцию. Подобные отношения можно констатировать у Ambulacralia и у позвоночных животных. У других животных с более простым ходом развития, как, например, у червей, моллюсков и многих членистоногих, безрезультатно искали подобные мезодермальные мешки; возможно, что у них вся мезодерма возникает без участия настоящих энтодермальных мешков, но может быть за счет эктодермы. В таких случаях мезодерма могла затем дифференцироваться на соматоплевру и спланхноплевру без мешковидной стадии. У Arthropoda с большим количеством питательного желтка мезодерма начинает функционировать очень рано, ее клетки пожирают желточные
* Die Polycladen (Fauna und Flora d. Golfes von Neapel), 1884, стр. 158.
237
зерна и перерабатывают их. Это делает понятным, что до последнего времени мезодерму часто принимали за энтодерму. В зависимости от фагоцитобласта, а отчасти от кинобласта возникает и рано (например, у Ctenophora) или поздно отделяется в качестве самостоятельного зародышевого листка мезодерма, играющая в эмбриональном развитии важную роль. В противоположность авторам, которые допускают возникновение мезодермы из половых органов или мышц, я думаю, что она как часть фагоцитобласта, подобно последнему, играла роль при захвате и резорбции пищи.
О значении мезодермальных фагоцитов в физиологических и патологических процессах, так же как о морфологии мезодермы, я высказывался в других работах* и, чтобы избежать повторений, сошлюсь на них.
В заключение я хочу еще заметить, что выяснение вопроса о происхождении многоклеточных необходимо для того, чтобы получить фундамент для сравнительной морфологии. Но до тех пор, однако, пока в разработке проблемы зародышевых листков выражено антигенеалогическое направление, решение важнейших вопросов будет наталкиваться на непреодолимые трудности 127. Поэтому я думаю, что при отсутствии положительных данных гипотетические построения могут считаться закономерными.
* Vergleichend-embryologische Studien, 3—5. Zeitschr. f. wiss. Zool., т. XXXVII, 1882, стр. 286, и т. XLII, 1885, стр. 648. — Untersuch. uber die intracellulare Verdauung bei Wirbellosen, Arbeiten des zool. Inst. d. Univ. zu Wien, т. V, 1883, стр. 147. — Untersuch. uber die mesodermalen Phagocyten einiger Wirbelthiere, Biol. Centralbl., т. III, 1883, стр. 560. — Ueber eine Sprosspilzkrankheit der Daphnien. Virchow's Archiv. т. XCVI, 1884, стр. 177. — Ueber die Beziehung der Phagocyten zu Milzbrandbacillen. Там же, т. XCVII, 1884, стр. 502.