
|
|
|||
|
АКАДЕМИЯ НАУК БЕЛОРУССКОЙ ССР
ИНСТИТУТ ФИЗИОЛОГИИ
Н. И. АРИНЧИН, Г. Ф. БОРИСЕВИЧ
МИКРОНАСОСНАЯ ДЕЯТЕЛЬНОСТЬ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ РАСТЯЖЕНИИ
|
|||||
|
МИНСК «НАУКА И ТЕХНИКА» 1986
|
|||||
|
УДК 612.741.61-06: 612.766.1
|
|||||||||
|
Аринчин Н. И., Борисевич Г. Ф. Микронасосная деятельность скелетных мышц при их растяжении.— Мн.: Наука и техника, 1986 —112 с.
Скелетные
мышцы — не только потребители крови, доставляемой сердцем, они обладают
самостоятельным присасывающе-нагнетатель-ным свойством,, которое
проявляется как при различных видах сокращения и массажа, -так и при
их растяжении. Обращается внимание на то, что.в повседневной
трудовой.деятельности, и. спорте должны совершенствоваться не только
сила, скорость и выносливость скелетных мышц, но и их микронасосная
деятельность с целью облегчения работы сердца, повышения его
надежности, укрепления здоровья и работоспособности человека.
Книга
предназначена для физиологов, патофизиологов, врачей, специалистов по
физической культуре и спорту, а также студентов биологического и
медицинского профиля.
Табл. 13. Ил. 20. Библиогр.: с. 97—111.
|
|||||||||
|
Научный редактор И. Р. Воронович, д-р мед. наук
Рецензенты:
В. В. Горбачев, д-р мед. наук, В. Н. Калюнов, д-р биол. наук
|
|||||||||
|
2007020000—136
А------------------------50—86
М316-86
© Издательство
|
|||||||||
|
ВВЕДЕНИЕ
|
||||||
|
Несмотря на
то что в последнее время все углубленнее и разностороннее становится
изучение этиопатогенеза заболеваний сердечно-сосудистой системы, они, к
сожалению, продолжают оставаться главной проблемой современной
медицины и занимают среди других заболеваний первое место в мире. В
связи с этим вопросы их профилактики и лечения привлекают внимание
многих исследователей. Одна из многочисленных причин широкого
распространения сердечно-сосудистых заболеваний— значительное понижение
двигательной активности человека. Предпринятые в этом аспекте
исследования с позиций вибрационной гипотезы кровоснабжения скелетных
мышц (Аринчин, 1960, 1961, 1966) привели к обнаружению их активной
внутриорганной присасывающе-нагне-тательной микронасосной функции
(Аринчин, Недвец-кая, 1973). На основании этого скелетная мышца
рассматривается как сложный сократительно-насосный орган, совмещающий
в себе одновременно выполнение^ не только физической работы, генерации
тепла и т. д., но и активное однонаправленное перекачивание крови из
артериального русла по внутримышечным капиллярам в венозное. Будучи
полностью изолированной, скелетная мышца способна осуществлять
замкнутую циркуляцию крови по искусственному кругу кровообращения,
вследствие чего она получила название внутримышечного периферического
«сердца».
Установлено,
что скелетные мышцы функционируют как присасывающе-нагнетательные
микронасосы при ритмических, тетанических сокращениях и даже в
условиях физиологического покоя (Аринчин, Недвецкая, 1974).
Общеизвестно,
что скелетные мышцы в организме человека и животных работают
координированно: в то.время как одни мышцы сокращаются, другие
растягиваются,
|
||||||
|
з
|
||||||
|
и наоборот,
т. е. растяжение и сокращение — элементы движения, протекающие
параллельно, согласованно, совокупность которых ведет к нормальному
двигательному эффекту. Представляло интерес выяснить, какую роль.в
гемодинамике играют скелетные мышцы при ритмическом их растяжении, а
также, как влияет физическая тренировка на взаимосвязь между
сердечно-сосудистой системой и деятельностью скелетных мышц в условиях
растяжения с точки зрения их присасывающе-нагнета-тельной способности.
В работе
представлены данные о том, что и в условиях растяжения изолированная
скелетная мышца функционирует как внутримышечное периферическое
«сердце». С этой целью использовались три методических подхода: 1.
Выявление нагнетательной способности скелетной мышцы и миокарда в
условиях растяжения. 2. Обнаружение присасывающего эффекта скелетной
мышцы при ритмическом растяжении. 3. Исследование способности
ритмически растягиваемой изолированной скелетной мышцы к
самостоятельному проталкиванию крови по искусственному кругу
кровообращения.
Полученные
результаты подтверждают существование присасывающе-нагнетательной
микронасосной функции в условиях ритмического растяжения изолированных
скелетных мышц, причем обнаружена ярко выраженная зависимость их
способности к насосной деятельности от силы растяжения, тонуса мышц,
физической тренировки и ряда других условий.
|
||||
|
Глава I
|
|||||||
|
ОБЗОР ЛИТЕРАТУРЫ
|
|||||||
|
Физиология
кровообращения, включающая центральное, регионарное, зональное и
другие разделы, остается еще мало разработанной. Эта проблема
привлекает внимание многих ученых: физиологов, патофизиологов,
морфологов, клиницистов и др. Литература по кровообращению настолько
велика, что охватить ее практически невозможно, поэтому ограничимся
рассмотрением только той, которая непосредственно относится к
интересующему нас вопросу.
ВНУТРИМЫШЕЧНОЕ ПЕРИФЕРИЧЕСКОЕ «СЕРДЦЕ», ЕГО ОБНАРУЖЕНИЕ И ХАРАКТЕРИСТИКА
Долгое время
считалось, что сердце — единственный гемодинамический насос, функция
которого заключается в активном проталкивании крови по большому и
малому кругу кровообращения.
Однако
прямые опыты измерения кровяного давления показали, что энергия сердца
почти целиком теряется в капиллярной сети. Эта энергия (99%) направлена
на преодоление сил трения и инерции, а также придание скорости току
крови в сосудистой системе, зависящих от числа форменных элементов,
вязкости крови, ангиоархи-тектоники и скорости кровотока в сосудах
(Schmid-Schonbein, 1975).
По данным
ряда авторов (Litter, Wood, 1954; Piper, 1958; Wiedeman, 1963; Folkow,
Mellander, 1964; Wood, 1965), основная масса крови содержится в области
низкого давления (70—80%): в капиллярах — 5% крови, в мелких артериях
— около 36% и наибольшее количество крови — в венулах и других мелких
венах органов. Сопротивление току крови в венах невелико. Наибольшее
сопротивление (80%) приходится на долю мелких сосудов: артериол,
прекапилляров, капилляров, артериове-
|
|||||||
|
5
|
|||||||
|
нозных анастомозов. Лишь 20% общего сопротивления составляет сопротивление в крупных артериях (Langen, 1961).
Для возврата
венозной крови к правому сердцу у человека в положении стоя требуется
давление, равное примерно 100 мм рт. ст., а энергии сердца для этого
явно недостаточно. Кроме того, при выполнении физической работы
мышечный, кровоток у среднего человека повышается с 800—1000 мл/мин до
15/20 л/мин при минутном объеме около 20—25 л/мин (Фолков, Нил, 1976),
что еще более остро ставит вопрос о трудностях венозного возврата к
сердцу. В связи с этим возникает предположение о том, что наряду с
сердцем в организме должны существовать какие-то дополнительные его
помощники.
В качестве
дополнительных механизмов, обеспечивающих продвижение крови к правому
сердцу, обнаружен ряд экстракардиальных факторов: «респираторный насос
Гендерсона», или присасывающая функция грудной клетки (Fleisch, 1930;
Прессман, 1952; Brecher, 1956; Knebel, Wick, 1958; Nordenstrom,
Norhagen, 1965; Schorer, 1966); дыхательные напряжения брюшного пресса
и диафрагмы (Alexander, 1951, 1963; Krug, Schlicher, 1960);
присасывающее действие желудочков сердца (Brecher, 1956; Bloom, 1956);
тонические сокращения вен (венопрессор-ный механизм), которые переводят
кинетическую энергию сердца в движение крови по сосудам и тем самым в
известной степени способствуют продвижению крови к сердцу (Кравков,
1916; Fleisch, 1920; Разенков, Иорданский, 1927; Golwitzer-Meiyer,
1932; Савицкий, 1936; Вальдман, 1940; Долго-Сабуров, 1958; Данилов,
1967 и др.); селезенка (Шор, 1925; Barcroft, 1926, 1932; Полосухин,
1957; Кричевская, Полосухин, 1966); венозная помпа или мышечный насос,
который благодаря сдавливанию сокращающимися скелетными мышцами
расположенных между ними венозных сосудов с клапанами направляет
кровь к сердцу (Hooker, 1911; Kjogh, 1922, 1970; Tiegerstedt, 1922:
Beecher, Field, Krogh, 1936; Seiro, 1938; Adams, John, 1939; Barcroft,
Dornhorst, 1949; Pollack, Wood, 1949; Warren, White, Belcher, 1949;
White, Warren, 1949; Аденский, 1953; Alexander, 1963; Guyton, 1963;
Folkow, Mellander, 1964; Gauer, Thron, 1965; Бабский и др., 1966;.
Фолков, Нил, 1976 и др.).
Однако многочисленными исследованиями доказано,
6
|
||||
|
что
экстракардиальные факторы кровообращения являются малоэффективными
помощниками сердца, а участие некоторых из них до сих пор оспаривается
и даже отрицается. Это относится к представлениям М. В. Яновского
(1909, 1922) о периферическом артериальном сердце, которое не нашло
подтверждения в физиологии и клинике (Френкель, 1935).
В связи с
этим вопрос о дополнительных помощниках сердца и в настоящее время
продолжает оставаться в центре внимания многих исследователей.
В свете
вибрационной гипотезы (Аринчин, 1960, 1966) было обращено внимание на
то, что скелетные мышцы в организме обладают
присасывающе-нагнетательной функцией, т. е. активно помогают сердцу в
продвижении крови по большому кругу кровообращения.
Насосная
функция скелетных мышц была обнаружена в опытах с искусственным
преграждением венозного оттока крови из сосудов икроножной мышцы
собаки. При раздражении мышцы ритмическими или тетаническими импульсами
застойное венозное давление не только достигало, но и достоверно
превышало уровень максимального артериального давления, развиваемого
сердцем (Аринчин, Недвецкая, 1973). Данные эксперименты проводились
при сохраненном артериальном притоке, т. е. кровь поступала в скелетную
мышцу под давлением, создаваемом сердцем и действием сил vis a tergo.
При отключении сил vis a tergo путем одновременного преграждения как
артериального притока, так и венозного оттока крови из сосудов мышцы и
ее сокращении (ритмическом или тетаническом) артериальное давление
падало вплоть до нуля, а венозное повышалось до 70—100 мм рт. ст. и
более.
Наиболее
убедительные доказательства в пользу того, что скелетная мышца
представляет собой активный гемо-динамический насос, действующий в
помощь сердцу, были получены на полностью изолированной икроножной
мышце (Аринчин, Недвецкая, 1974, 1976; Аринчин, Недвецкая, Кабуш,
1975). Показано, что при подключении изолированной икроножной мышцы к
искусственному кругу кровообращения и одновременном ее раздражении
импульсами различной частоты кровь продвигалась из внутримышечных
капилляров в бедренную вену, а из нее по искусственному кругу в
бедренную артерию и возвращалась в капилляры. Следовательно, мышца
обладает
|
|||||
|
7
|
|||||
|
способностью
самостоятельно осуществлять замкнутую циркуляцию заключенной в ней
крови по образу и подобию сердца, вследствие чего получила название
периферического «сердца».
При создании
ретроградного кровотока путем соединения Т-образными стеклянными
канюлями артерии с веной, а вены с артерией (в результате чего
артериальная кровь направлялась в вену) обнаружено, что насосная
функция скелетных мышц отсутствует, т. е. кровоток может осуществляться
только однонаправленно — из артерий по капиллярам в вены. Вероятно,
однонаправленность движения крови связана не только с наличием
клапанов в венах, но и в большей мере с однонаправленной
нагнетательной микронасосной деятельностью скелетных мышц.
Электронномикроскопическими
исследованиями установлено, что клапаны находятся в тончайших венах и
даже посткапиллярных венулах диаметром меньше 0,05—10 мкм (Miani,
Ruberti, 1958; Staubesand, Rulffs, 1958; Куприянов, 1969 и др.). Вопрос
об участии клапанов в венозном кровообращении решается многими
авторами противоречиво. Одни авторы придавали большое значение роли
клапанов в движении крови по венам (Hooker, 1911; Seiro, 1938; Adams,
1939; Guyton, 1963; Folkow, Mellander, 1964; Бабский и др., 1966;
Kxogh, 1970). Другие же считали, что венозные клапаны в деятельности
венозной помпы играют очень незначительную роль, а некоторые из них
полагали, что даже при отсутствии клапанов может быть улучшение
венозного возврата крови во время ритмических мышечных движений
(Alexander, 1951; Brechei, 1956).
Все это
относится к крупным венам, расположенным между мышцами, а
функциональное значение микроклапанов в венозной системе самих
скелетных мышц до сих пор не изучено. По мнению В. В. Куприянова
(1969), микроклапаны вряд ли могут так же, как и клапаны крупных вен,
удерживать кровь, перемещающуюся против силы тяжести, поскольку
разница в венозном давлении здесь ничтожна, а число самих
микроклапанов невелико.
Одним из
факторов, влияющих на приток крови к сердцу из вен, является
деятельность скелетных мышц, так называемая венозная помпа. Еще У.
Гарвей (1628) заметил, что сокращающиеся скелетные мышцы сдавли-
8
|
||||
|
в а ют
тонкостенные вены с клапанами, проходящие между мышцами и тем самым
способствуют выталкиванию крови в сторону сердца. Но данный факт может
иметь место только при мышечных сокращениях, чередующих-ся с
расслаблением. При длительном тетаническом сокращении мышц, не
прерываемом расслаблениями, мышечные волокна надолго пережимают
сосуды, и тогда скелетные мышцы не играют роль мышечного насоса. В
этом заключается одно из различий между венозной помпой (или мышечным
насосом) и внутримышечным периферическим «сердцем». Если даже венозная
помпа функционирует в самой мышце, то, как показано исследователями,
занимающимися ею, она проявляется только при ритмических сокращениях.
Но из данных литературы известно, что при длительном статическом
напряжении мышцы кровоток через нее может не только не изменяться, а
даже возрастать (Тхоревский, 1973 и др.). Это позволяет предположить,
что наряду с венозной помпой в мышце имеется какой-то дополнительный
механизм, который проявляется и при ритмических, и при тетаниче-ских
сокращениях (наличие клапанов во внутримышечных сосудах не
обязательно, поскольку этот механизм существует в основном на уровне
капилляров).
Присасывающий
эффект микронасосной деятельности скелетных мышц наиболее ярко
проявился в серии экспериментов с применением прямого ' ручного
массажа мышцы. В данном случае при одновременном пережатии сосудов
икроножной мышцы застойное венозное давление резко возрастало,
достигая 300 мм рт. ст., а артериальное падало до нуля и даже
становилось отрицательным. Массаж может рассматриваться как
своеобразная модель межмышечных взаимоотношений, при которых сокращения
близлежащих мышц, взаимодействующих друг с другом, приводят к
значительному повышению эффективности их насосной функции.
Поперечно-полосатая
мускулатура животных и человека проявляет определенную активность и в
сострянии физиологического покоя, за счет чего поддерживается тонус
мышечной ткани и выполняется антигравитационная работа (Верещагин,
Жуков, Иванов, 1947; Gopfert, 1952; Gopfert, Eiff, Howind, 1953;
Юсевич, 1953, 1963; Жуков, 1956, 1959, 1964, 1976; Кесарева, 1960;
Панфилов, 1964; Buchtal, Rosenfalk, 1966; Водолазский, 1966 и др.).
Тонус мышцы, по мнению Р. С. Персон (1969), услов-
|
|||||
|
9
|
|||||
|
но делится
на 3 категории явлений: 1—состояние мышцы .(ее твердость, упругость); 2
— некоторый уровень возбуждения мышцы, обусловленный тем, что часть
а-мото-нейронов ритмически разряжается с относительно небольшой
частотой; 3 — некоторый уровень возбудимости рефлекторных механизмов,
регулирующих состояние мышц.
В скелетной
мускулатуре позвоночных животных имеются 3 типа мышечных волокон:
нетонические быстрые, характеризующиеся быстрыми фазными сокращениями и
быстрой утомляемостью; тонические, медленно сокращающиеся,
отличающиеся низкой возбудимостью и малой утомляемостью; переходные,
обладающие промежуточными свойствами (Лебедева, 1930; Kruger, 1952;
Кол-лос, 1957; Жуков, 1964, 1969; Махмудов, 1969; Магазаник, Наследова,
1974 и др.).
А. Махмудов
(1969) считал, что тонические мышечные волокна благодаря специфике
строения и биохимическим, физиологическим особенностям работают не
одновременно, а попеременно, сменяя друг друга, что и обеспечивает
длительное, без утомления, сокращение мышцы в целом.
Таким
образом, тонус — это не сопровождающееся значительным усилением обмена
веществ, продолжительное по времени сокращение мышц, отличающееся
большой экономичностью.
При изучении
дрожательного паралича у паркинсони-ков была открыта микровибрация
тела. Основной причиной микровибрации организма при полном видимом
покое является вибрация поперечно-полосатой мускулатуры, т. е.
сокращения отдельных мышечных волокон или небольших их групп,
совершающиеся в определенной последовательности, что свойственно всем
теплокровным животным (Rohracher, 1955, 1959).
Н. Rohracher
(1959) показал, что между мышечным тонусом и микровибрацией существует
связь, вытекающая из их основных свойств. Эта взаимосвязь заключается
в том, что постоянно совершающиеся сокращения отдельных двигательных
единиц (причина микровибрации) лежат в основе мышечного тонуса.
Мышечный тонус—это мышечное напряжение, возникающее на основе такого
сокращения. Таким образом, тонус и микровибрация — различные действия
одного и того же мышечного процесса.
Для выяснения роли тонического напряжения скелет-
|
|||||
|
ю
|
|||||
|
ных мышц во
внутримышечной гемодинамике проведены эксперименты с внутриартериальным
введением препарата кураре, который блокирует передачу энергии с нерва
на мышечное волокно и тем самым значительно снижает тонус мышцы
(Аринчин, Недвецкая, 1974).
При
преграждении венозного оттока крови из сосудов икроножной мышцы,
находящейся при этом в покое, застойное венозное давление повышалось в
меньшей степени вследствие выключения части микронасосов в скелетной
мышце. Это говорит о том, что микронасосы работают и в покое, в
некураризированной мышце. Уменьшение повышения застойного венозного
давления, а не полное его отсутствие при кураризации мышцы объясняется
тем, что микровибрация при введении кураре снижается, но никогда не
исчезает полностью (Иванов, 1965), сохраняется и перфузия мышцы.
На
микронасосную деятельность скелетных мышц в условиях физиологического
покоя указывают также данные о том, что появление видимой фибрилляции
мышечных волокон способствует дальнейшему повышению застойного
венозного давления.
Многие
исследователи обращали внимание на участие мышечного тонуса в
гемодинамике организма (Beiglbock, Junk, 1937; Henderson, 1938;
Иванова, 1960; Новиков, . 1966; Панфилов, 1968 и др.). Они считали, что
внутримышечное давление скелетных мышц является одним из
экстракардиальных факторов кровообращения, за счет которого
поддерживается более высокое капиллярное давление, увеличиваются
скорость кровотока, а также объем крови, протекающей в единицу времени,
при сниженном периферическом сопротивлении сосудистой сети и
артериальном давлении. Следовательно, по их мнению, при высоком тонусе
скелетных мышц работа сердца протекает в более благоприятных условиях
и более эффективно, чем при пониженном тонусе.
Механизм
деятельности скелетных мышц как мощного гемодинамического насоса можно
объяснить с позиций вибрационной гипотезы (Аринчин, 1961). Сущность
этой гипотезы заключается в том, что мышечные волок- ■ на, асинхронно
сокращаясь, ритмически пережимают кровеносные сосуды, расположенные в
мышце, и тем самым способствуют продвижению крови в сторону меньшего
давления, развивая при этом присасывающе-нагнета-тельный эффект. Эта
гипотеза основана на многочислен-
|
|||||
|
11
|
|||||
|
ных,
полученных ранее, данных о том, что в основе мышечного сокращения
лежат асинхронные сокращения отдельных двигательных единиц (Wachholder,
1931; Bou-man, Rijnberk, 1938; Brandstater, 1938 и др.).
Среди
разнообразных видов двигательной активности животных и человека большую
роль играет динамическая и статическая работа, гармоническое сочетание
двух типов сокращения скелетных мышц — тонического и те-танического.
Существуют
два различных взгляда на природу тонуса и тетануса поперечно-полосатых
мышц. Первая точка зрения предполагает два различных анатомических
субстрата, один-из которых специализирован на функцию тетануса, а
второй — на функцию тонуса (Верещагин, Жуков, Иванов, 1947; Жуков,
Попова, 1957). Тетаниче-ский и тонический аппараты могут получать
раздельную иннервацию со стороны соматической нервной системы.
Тетанические и тонические нервные волокна отличаются друг от друга по
ряду признаков, в частности по месту выхода из спинного мозга. Вторая
точка зрения заключается в том, что один и тот же субстрат в
зависимости от потребности организма может функционально
перестраиваться на тоническую или тетаническую деятельность. Считают,
что тонические сокращения скелетной мышцы по своему механизму не что
иное, как разновидность слабого тетануса.
Таким
образом, в основе динамической или статической работы лежит
тетаническое сокращение мышцы, образующееся путем пространственной или
временной суммации одиночных сокращений, ибо, как правило, нервные
импульсы приходят к фазным мышечным волокнам не одновременно, а
асинхронно. При сокращении мышцы происходят асинхронные сокращения
больших или малых групп мотонейронов, т. е. сокращение целой мышцы
является суперпозицией актов сокращения составляющих мышцу
двигательных единиц. Двигательные единицы «включаются» асинхронно по
программе, заложенной в управляющем нервном приборе, и лишь при
предельных мышечных усилиях мотонейроны, иннерви-рующие мышцу, начинают
разряжаться синхронно (Ма-газаник, Наследова, 1974).
Взаимосвязь
между мышечным усилием и биоэлектрической активностью была установлена
давно (Введенский, 1951, 1952; Wachhoider, Altenburger, 1926;
|
|||||
|
12
|
|||||
|
Wachholder,
1931; Киселев, Маршак, 1935; Персон, 1965 1969). Но одни авторы
(Кожевников и др., 1961) считали] что между электрическим и
механическим эффектом мышцы существует линейная зависимость, а другие
(Гур-финкель, Код, Шик, 1965) отмечали отклонения от прямолинейной
зависимости при больших нагрузках.
Такая же
зависимость существует между частотой звуковых осцилляции и колебаниями
мышечных волокон (Bouman, Rijnberk, 1938; Brandstater, 1938).
Методом
синхронной регистрации электромиограм-мы, фономиограммы, объемного
пульса, объемно-пульсового кровоснабжения, венозного, артериального
давлений и тонуса сосудов методом ангиотензиотонографии, объемной
скорости кровотока методом окклюзии, тонического напряжения мышц и
других показателей была выявлена зависимость кровоснабжения мышц
предплечья от вибрационных колебаний мышечных волокон при различных
произвольных статических напряжениях (Ермолова, 1970, 1971; Аринчин,
Ермолова, 1971). Показано, что амплитуда осцилляции электромиограммы
изменялась пропорционально величине статических усилий, частота
осцилляции фономиограммы в первую половину статической нагрузки
изменялась в зависимости от величины усилий, затем оставалась на
повышенном уровне. Амплитуда электромиограммы линейно зависела от
величины статического усилия. Внутримышечные сосуды даже при
максимальном напряжении не пережимались полностью, ибо объемная
скорость кровотока в предплечье спортсменов в этих условиях
увеличивалась на 20—24% по сравнению с покоем.
Ряд авторов
отмечали,, что даже в покоящейся мышце внутримышечное давление выше,
чем внутрисосудистое в капиллярах и венах, а в сокращенной — оно еще
выше как при динамической, так и при статической работе. Это
превосходящее в несколько раз давление должно было бы перекрыть сосуды,
расположенные в мышце, чего на самом деле не происходит (Sylvest, Hvid,
1959; Аринчин, 1960, 1961).
Многими
исследователями показано, что при статических нагрузках, когда
внутримышечное давление намного превышает внутрисосудистое, кровоток в
мышце ухудшался, но все же сохранялся, хотя и на более низком уровне
(Меньшикова, 1938; Могендович, 1957) или незначительно (Clarke, Hellon,
Lind, 1958), или довольно
|
|||||
|
13
|
|||||
|
существенно
увеличивался (Шабунин, 1959, 1965; Арин-чин, 1961; Humphreys, Lind,
1963; Титиевская, 1964; Ве-vegard, Shepherd, 1967; Скардс, Шмульян,
1968; Тхорев-ский, 1968; Шмульян, 1968; Озолинь, Порцик, 1970).
P. W.
Humphreys, A. R. Lind (1963) методом плетизмографии в сочетании с
анализом изменений систолического артериального давления и сокращения
отдельных мышц предплечья обнаружили, что во время статических
сокращений с силой 10—70% максимального произвольного напряжения
кровоток в предплечье увеличивался до 10—40 мл/100 см3
ткани. Они полагали, что увеличивающееся во время сокращения
внутримышечное давление, хотя и повышает сопротивление кровотоку, но
не может перекрыть его, пока сила произвольного напряжения не превысит
70% максимального произвольного напряжения.
Принимая во
внимание данные, полученные А. С. Ермоловой, увеличение
внутримышечного кровотока во время статических сокращений можно
объяснить тем, что кровоснабжение скелетных мышц в этих условиях
поддерживается за счет асинхронных сокращений мышечных волокон,
воздействующих на кровеносные сосуды.
Результаты
А. С. Ермоловой подтверждены экспериментами с синхронной регистрацией
ангиотензиотоно-граммы, объемной скорости кровотока, электромиограм-мы,
фономиограммы, вибромиограммы изолированной икроножной мышцы собаки.
Объемная скорость кровотока при раздражении мышцы импульсами различной
частоты увеличивалась синхронно с повышением ее электрической,
звуковой, механической активности (Аринчин, Ермолова, Наймитенко,
Недвецкая, 1973). Мышечные волокна,- асинхронно сокращаясь со звуковой
частотой, способствуют увеличению кровотока во время статических
нагрузок (Аринчин, Кулаго, 1969; Наймитенко, 1970, 1971, 1972; Аринчин,
Ермолова, Наймитенко, Недвецкая, 1972).
Л. В.
Наймитенко (1971) провел серию опытов по наливке рентгеноконтрастным
веществом сосудов икроножной мышцы собаки, находящейся в состоянии
покоя, под воздействием электрической стимуляции, при ее прямом массаже
и внешней вибрации. В результате этих исследований обнаружено, что под
влиянием механических воздействий мышечных волокон на сосуды во время
сокращения, массажа, внешней вибрации частицы раство-
|
|||||
|
14
|
|||||
|
pa проникали
через капилляры в бедренную вену, о чем свидетельствовала более
разветвленная и густая сеть сосудов по сравнению с контролем и
заполнением контрастным веществом вены.
При изучении
методом микрокиносъемки кровотока во внутримышечных сосудах
поперечно-полосатых и гладких мышц в динамике получены результаты,
подтверждающие основные положения вибрационной гипотезы (Аринчин,
Недвецкая, Наймитенко, Новгородцев, 1972; Наймитенко, 1972). W.
Spalteholtz (1888), изучая артерии, вены, капилляры мышц животных и
новорожденных детей, первым точно описал расположение кровеносных
капилляров в мышцах. A. Kjogh (1927) описал «первичную» и «вторичную»
сеть капилляров. Рядом авторов было отмечено, что между числом
капилляров в мышечных пучках и толщиной волокон, а также величиной
энергетических затрат мышцы существует прямая зависимость (Жданов,
1964; Иванова, 1968; Маркизов, 1969). Формирование мышечных капилляров
происходит параллельно росту животного: при удлинении мышечных
волокон увеличивается число капилляров вдоль каждого мышечного
волокна и удлиняются уже образовавшиеся капилляры. Согласно данным Lee
(1958), капилляры в быстросокращающихся мышцах прямолинейны, а в
медленных извилисты.
В мышцах,
состоящих из поперечно-полосатой ткани, сосуды, в том числе и
капилляры, располагаются, параллельно мышечным пучкам и волокнам
(Привес, 1968, Иванова, 1973; Шошенко, 1973 и др.).
М. J.
Plyiey, А. С. Groone (1975) показали, что одно мышечное волокно
окружают от 0 до 9 капилляров. Среднее количество волокон, приходящихся
на 1 капилляр, колебалось от 2 до 3,2.
В мышцах
капилляр от капилляра расположен на расстоянии от 11 (Krogh, 1927) до
50 мк (Zweifach, 1959) и проходит в местах схождения двух или
нескольких граней волокон.
Гистологические
исследования показали, что нервно-мышечный синапс чаще всего находится
в центральной части волокна (Desmedt, 1958; Coers, Woolff, 1959;
Chris-tensen, 1959).
На небольшом
участке мышечного волокна В. П. Ги-лев (1961) обнаружил все стадии
сокращения саркоме-ров. Оказалось, что миофибриллы, находящиеся дальше
|
|||||
|
15
|
|||||
|
от места
раздражения, менее сокращены, и наоборот, миофибриллы, расположенные
ближе к месту раздражения, сильно сокращены. Между этими миофибриллами
располагаются остальные, на которых можно наблюдать все промежуточные
стадии сокращения.
На основании
этого было высказано предположение, что при сокращении мышцы длина
капилляра и его диаметр под возросшим внешним давлением станут меньше,
объем капилляра уменьшится и содержащаяся в нем кровь вытолкнется. При
расслаблении мышечных волокон объем капилляра увеличивается, и он снова
заполнится кровью (Аринчин, Ермолова, Наймитенко, Недвецкая, 1972;
Аринчин, Недвецкая, 1974).
Исследованиями,
проведенными в лаборатории кровообращения Института физиологии АН
БССР, обнаружено, что микронасосная функция скелетных мышц усиливается
при общей гипотермии организма, в случае возникновения холодовой
мышечной дрожи (Аринчин, Недвецкая, Борисевич, 1975, 1977). Это
объясняется тем, что мышечная дрожь обусловлена повышением
сократительной способности мышечных волокон или небольших их групп, т.
е. сократительной способности мышц (Бартон, Эдхолм, 1957; Иванов, 1965
и др.).
Таким
образом, скелетные мышцы представляют собой своеобразные
внутримышечные периферические «сердца», которые функционируют на
внутриорганном уровне при любых проявлениях жизнедеятельности
организма как при динамической, так и при статической работе.
Внутримышечные
периферические «сердца» действуют в области прекапилляров, венул, вен,
заключенных внутри мышцы, активно перекачивая кровь из артерий по
капиллярам в вены.
В последнее
время показано, что "и миокард в силу его значительного
анатомо-морфологического и функционального сходства со скелетными
мышцами обладает активной внутримиокардиальной насосной функцией,
которая заключается в том, что в условиях прямого массажа,
ритмического растяжения и естественных сокращений сердца, кровь
благодаря сократительной способности миокарда перекачивается из
коронарных артерий по капиллярам в венозные коллекторы сердца (Аринчин,
Недвецкая, 1976).
Значение внутриорганной микронасосной способности
|
|||||
|
16
|
|||||
|
скелетных
мышц становится еще более очевидным, если лринять во внимание то, что
на долю скелетных мышц приходится до 50% массы тела человека
(Васильева, Степочкина, 1968) и они обильно (2000—3000 капилляров в 1
мм2) кровсснабжаются (Krogh, 1927).
Следовательно,
в аппарате кровообращения существует не один, а несколько насосов,
составляющих систему гемодинамических насосов организма: среди них
первый уровень принадлежит сердцу, второй — внутримышечным
периферическим «сердцам», третий — венозным помпам. Первый и третий
уровни изучены достаточно хорошо, тогда как второй до недавнего
времени оставался неизвестным.
Ранее
проведенными исследованиями установлено, что скелетные мышцы обладают
присасывающе-нагнета-тельным свойством при различных видах сокращения,
массажа, холодовой мышечной дрожи, но как ведет себя мышца при
растяжении в условиях реципрокной деятельности мышц-антагонистов
предстояло всесторонне изучить.
РАСТЯЖЕНИЕ КАК ФУНКЦИОНАЛЬНОЕ СОСТОЯНИЕ СКЕЛЕТНЫХ МЫШЦ
Двигательная
деятельность в естественных условиях представляет собой сочетание
статической и динамической работы, которая осуществляется на фоне
тонического напряжения скелетных мышц.
Элементарная координация работы скелетных мышц-антагонистов имеет в своей основе механизм реципрокной их иннервации.
Однако
типичных мышц-антагонистов мало, обычно мышцы являются антагонистами
для конкретной ситуации, при ее же изменении они находятся в других
соотношениях. Изучение биотоков антагонистических мышц у человека при
волевых движениях показало, что возможны как реципрокные отношения
этих мышц, так и их одновременное сокращение или расслабление (Wagner,
1925; Wachholder, 1928; Персон, 1965 и др.).
Вопросами
взаимоотношений мышц-антагонистов занимались многие исследователи
(Введенский, 1951, 1952; Scherrington, Hering, 1897; Введенский,
Ухтомский, 1952; Кекчеев, 1946; Уфлянд, 1952, 1956; Гурфинкель, 1953;
Донская, 1956; Границ, 1957; Фарфель, 1960; Захарьянц, 1963; Беккер,
1965; Персон, 1965; Костюк, 1968 и др.).
|
|||||||
|
2. Зак. 837
|
|||||||
|
17
|
|||||||
|
В основе
реципрокных отношений лежат ироприоцеп-тивные рефлексы, принимающие
участие как в поддержании мышечного тонуса, так и в обеспечении
координации движений. Проприоцептивные рефлексы — рефлексы с обратной
связью по функциональной структуре и рефлексы растяжения по
происхождению.
Функцию
собственно' проприорецепторов, воспринимающих растяжение и сокращение
мышцы, выполняют веретенообразные клетки, расположенные в ней, что
было экспериментально доказано путем перерезки спинномозговых
корешков у кошек, обезьян, лягушек.
В настоящее время выделены а-, у-мотонейроны,
воздействующие на возбудимость проприорецепторов. Импульсы, идущие к
мышечным веретенам по у-эфферен-там, сказываются на степени их
напряжения, в результате чего изменяется их чувствительность к
напряжению мышцы (Tiegs, 1953). На проприорецепторы оказывают
непрерывное влияние надсегментарные отделы мозга. Они также тонко
реагируют на различные адекватные воздействия со стороны скелетных
мышц. Это проявляется в рефлекторном повышении тонуса и в колебании
возбудимости мышц как на стороне раздражения, так и на симметричных
мышцах противоположной стороны.
Таким
образом, реципрокные отношения складываются еще в спинном мозгу, а
контролируются высшими отделами центральной нервной системы.
Следовательно,
элементы движения представляют собой совокупность процессов растяжения
и сокращения, которые осуществляются параллельно, координированно и при
своем взаимодействии ведут к нормальному биологическому двигательному
акту.
Большое
участие растяжение принимает при выполнении циклических движений,
которые являются основой наиболее распространенных видов локомоций.
Известно,
что скелетные мышцы обладают упругими и эластичными свойствами, в
значительной мере способствующими растяжению и сокращению мышцы в
естественных условиях. Однако упругость мышц несовершенна. Е. К- Жуков
(1959) считал, что при растяжении выделяются две фазы: 1 — быстрое
растяжение мышцы, которое отражает деформацию, упругих компонентов; 2 —
медленное ее растяжение, свидетельствующее о наличии в мышце
внутреннего трения или вязкости. Значит, в начале
|
||||
|
18
|
||||
|
растяжения
мышца оказывает незначительное сопротивление растягиваемой силе, при
дальнейшем растяжении сопротивление мышцы растягиваемой силе растет.
A. Haman et
al. (1974) также обратили внимание на нелинейность растяжения мышцы,
так как изменение ее длины не пропорционально напряжению. По полученным
ими данным, мышца может растянуться свыше 150% своей длины в покое.
Нелинейность
растяжения мышцы, по всей вероятности, зависит от того, что в самой
мышце одни ее участки сокращаются, в то время как другие, находящиеся
еще в покое, растягиваются. Следовательно, мышцы наряду с сократимостью
обладают и свойством растяжимости. Это свойство можно наблюдать на
изолированной мышце, находящейся в покое, если растягивать ее грузом.
S. Gollaunaugh, M. Brookes (1974), исследуя влияние натяжения т. tibialis anterior при
нагрузке от 0 до 200 г на длину мышечных волокон, обнаружили, что
кривая зависимости логарифма среднего процента изменения длины от
величины натяжения имеет экспоненциальный-характер с тремя перегибами,
соответствующими трем фазам состояния мышцы. Во время первой фазы
наблюдается скольжение мышечных пучков, обусловленное наличием
эластичных волокон в перимизиуме. Во время второй фазы филаменты актина
и миозина разделены, скорость удлинения снижается, хотя сила растяжения
продолжает возрастать. Это происходит до тех пор, пока не наступит
третья фаза, когда тонкие спиральные колла-геновые волокна,
расположенные в эндомизиуме, не вытянутся параллельно миофибриллам.
Такую зависимость авторы нашли как для напряженной, так и для
расслабленной мышцы.
Общепринятым
считалось, что дальше миофибриллы не могут удлиняться без разрушения,
ссылаясь на теорию скольжения, согласно которой перерастяжение мышцы
приводит к потере миофибриллярной способности сокращаться. Однако
исследованиями Н, В. Самосудовой, Г. М. Фра нка (1962), Г. М. Франка
(1970) показано, что сокращение все же имеет место.
Как для
сокращения, так и для растяжения прото-фибрилл необходима АТФ. Под
влиянием ее резко возрастает пластичность актомиозиновых нитей: под
нагрузкой они сильно растягиваются, а в свободном состоянии
сокращаются.
|
|||||
|
19
|
|||||
|
Н. Е.
Huxley, J. Hanson (1954) с помощью электронного микроскопа установили,
что при растяжении мышечных волокон удлинение саркомера происходит за
счет «вытягивания» тонких протофибрилл из промежутков между толстыми, а
при сокращении, наоборот, происходит их «вдвигание».
По
современным представлениям, при растяжении мышцы увеличивается степень
взаимодействия актино-вых и миозиновых нитей, а так как
взаиморасположение актиновых и миозиновых нитей при растяжении
определяет их взаимодействие при сокращении, то сила сокращения мышцы
увеличивается.
Таким
.образом, растяжение представляет собой процесс, противоположный
сокращению, и является необходимым условием для нормального сокращения
и в целом функционирования всей мышечной ткани.
Многочисленными
работами показано, что растяжение влияет на функциональные свойства
скелетной мышцы (Бауэр, Барздыко, 1936; Стома, 1956, 1958; Кондраев,
Чуваев, Чуваева, 1962; Гоциридзе, 1968; Глазырина, 1969; Ковшова, 1972
и др.).
Установлено,
что умеренное натяжение мышцы, как •изолированной, так и в условиях
целостного организма, меняет ряд ее свойств: увеличивается высота
одиночного сокращения, укорачивается скрытый период сокращения (Мужеев,
1935), уменьшается реобаза (Стома, 1956, 1958), увеличивается скорость
возникновения возбуждения (Стома, 1956, 1958; Гоциридзе, 1968).
По данным
ряда авторов, реакция на растяжение является частным случаем
парабиотического процесса (Га-нелина, 1962; Глазырина, 1969).
П. В.
Глазырина (1969) обнаружила, что при умеренном растяжении мышцы
возбудимость и сократительная способность мышечного волокна повышаются,
а чувствительность к избытку ионов калия понижается; при сильном
растяжении, наоборот, происходит падение возбудимости и сократительной
способности, в мышце нарастают паранекротические изменения. Имеются
данные, свидетельствующие о том, что при средней силе растяжения в
мышце усиливаются обменные процессы (Ганелина, 1962).
Реобаза и
хронаксия растягиваемой мышцы изменяются в зависимости от величины
нагрузки, а при длительном растяжении М. Ф. Стома (1956) наблюдал
трехфаз-
|
|||||
|
20
|
|||||
|
ность
изменения функционального- состояния мышцы: во время первой фазы
происходит кратковременное увеличение хронаксии, во время второй —
укорочение хронак-сии и увеличение реобазы, а в третьей фазе —
удлинение хронаксии и увеличение реобазы. Прекращение' растяжения
вызывает постепенное повышение лабильности и возбудимости мышцы до
исходных величин, которое сменяется вторичной волной понижения тех же
показателей. Характеризуя работоспособность утомленной мышцы, Ф. Е. Тур
(1938), М. В. Кирзон (1959), Н. Н. Лебедева (1956) обнаружили, что при
дополнительном растяжении работоспособность мышцы увеличивается. Это,
по мнению М. В. Кирзон (1959), объясняется тем, что добавочное
растяжение дополнительно включает в работу новые • нейромоторные
единицы.
Однако по
вопросу о механизме действия растяжения на скелетные мышцы нет единого
мнения. Одни авторы считают, что растяжение непосредственно действует
на мышечное волокно, изменяя его динамические свойства (Введенский,
1893 и др.). Другие предполагают, "что растяжение воздействует на
нервно-мышечный аппарат и тем самым улучшает передачу возбуждения с
нерва на мышечное волокно (Кирзон, 1959; Сагдеев, 1971).
При
раздражении мышцы ускорение нервно-мышечной передачи возбуждения
вызывается за счет увеличения освобождения ацетилхолина из нервных
окончаний, что ведет к возрастанию амплитуды потенциалов концевых
пластинок. Вовлечению новых волокон в активность отчасти способствует и
повышение их возбудимости (Сагдеев, 1971).
Экспериментально
на кураризированном нервно-мышечном препарате лягушки Т. Н. Ониани, А.
А. Унгиадзе (1964) показали, что растяжение мышцы вызывает увеличение
амплитуды потенциала концевых пластинок, а также повышение
возбудимости, мышечных волокон, что облегчает передачу возбуждения с
нерва на мышцу.
Третьи,
составляющие большинство исследователей, получили многочисленные
доказательства о том, что растяжение в условиях целостного организма
является специфическим раздражителем проприорецепторов и тем самым
воздействует на центральную нервную систему с последующим рефлекторным
влиянием на нервно-мышечный аппарат (Стома, 1956; Уфлянд, 1956;
Гранит, 1957; Стома, Барвитенко, 1964; Григорьев, 1972 и др.).
2Е
|
||||
|
Еще с конца
прошлого века известно, что всякое изменение длины или напряжения
мышцы раздражает заключенные в ней проприорецепторы. В ее
'экстрафузаль-ных волокнах появляются рефлекторные токи действия.
Детальное изучение этого вида мышечных рефлексов было проведено Ч.
Шеррингтоном с сотр. (1935).
Раздражение
мышечных рецепторов при растяжении ведет к рефлекторному повышению
сократимости мышц. При изучении электрической импульсации, возникающей
в чувствительных окончаниях мышечного веретена лягушки в условиях
растяжения, обнаружены линейная зависимость между частотой импульсов и
величиной абсолютного удлинения мышцы и логарифмическая зависимость
между растягивающей силой и абсолютным ее удлинением (Заранов,
Метревели, Лежава, 1966).
При
ритмическом механическом раздражении про-приорецепторов в них
появляются рефлекторные биотоки или пачки биотоков, синхронные с
частотой раздражения. Вследствие неоднородности нейромоторных единиц и
соответственно неодинаковой скорости проведения импульсов по
афферентной и эфферентной части проприо-цептивной дуги в ответ на
раздражение возникают не отдельные биотоки, а пачки биотоков
(Григорьев, 1972).
Обнаружено,
что ступенчатое растяжение икроножной мышцы крыс приводит к
возбуждению сначала единиц с малой амплитудой разрядов, а по мере
усиления растяжения увеличивается общее количество работающих
одновременно единиц, среди которых увеличивается число единиц,
генерирующих разряды большой частоты (Бурсиан, 1972).
А. С.
Степанов, Б. М. Дементьев (1967) установили, что растяжение или
сокращение оказывает влияние не только на проприорецепторы мышц, но и
непосредственно на мышечные волокна. Причем насильственное растяжение
развивает в мышце напряжение примерно на 1/5 часть больше по сравнению
с изометрическим сокращением, вызванным электрическим раздражением
мышцы. - Полезная работа, выполняемая непосредственно после растяжения,
может быть значительно больше, чем работа той же мышцы при укорочении
в аналогичных прочих условиях, но из состояния изометрического
сокращения, т. е. без растяжения. Это связано как с большей
сократительной способностью мышцы, развиваемой после
22
|
||||||
|
растяжения (Стома, 1956; Кирзон, 1959; Глазырина, 1969 и др.), так и накоплением эластической энергии (Cavag-па, 1973).
В основном
механическое растяжение мышцы использовалось как адекватный
раздражитель проприорецепто-ров мышечной ткани и с этой целью
применялось при изучении проблемы рецепции и рефлекторной
деятельности. Однако многочисленные данные свидетельствуют о том, что
раздражение проприорецепторов оказывает не только координирующее
влияние на локомоторный аппарат, но и весьма существенно влияет на
сердечно-сосудистую систему по механизму моторно-висцеральных
рефлексов (Вазилло, Могендович, 1961; Могендович, 1957, 1962, 1968,
1971, 1972).
Ряд авторов,
занимаясь этим вопросом, обнаружили, что растяжение, сокращение и даже
незначительное изменение тонуса скелетных мышц, воздействуя на
про-приорецепторы, приводят у животных к различным эффектам как со
стороны работы сердца, так и сопротивления сосудов различных частей
тела.
В острых
опытах на кошках Е. А. Квасова, П. А. Некрасов (1951) получили
прессорный эффект при раздражении четырехглавой мышцы бедра, который
исчезал после перерезки задних корешков спинного мозга. Аналогичные
результаты получил В. П. Колычев (1959), который в качестве
экспериментального объекта использовал бульбоспинальных лягушек. ;
Депрессорные
реакции кровяного давления с последующей гипертензией при растяжении
мышцы кошки грузом были описаны S. Scoglung (1960).
Показано,-
что при раздражении проприорецепторов путем растяжения происходит
рефлекторное сужение кровеносных сосудов, в связи с чем повышается
общее кровяное давление. Особенно большая роль в повышении
артериального давления приписывается рефлекторным изменениям просвета
кровеносных сосудов внутренних органов (Георгиев, 1961).
При изучении
рефлекторного влияния раздражения проприорецепторов на деятельность
сердца получены противоречивые результаты. У холоднокровных животных
наблюдалось рефлекторное увеличение амплитуды и частоты сердцебиений
(Бельтюков, Могендович, 1947; Глебовский, 1949), а у теплокровных при
раздражении проприорецепторов путем растяжения не было обнаружено
|
|||||
|
23
|
|||||
|
изменений в деятельности сердца (Квасов, Науменко,' 1951; Георгиев, 1961).
По мнению М.
Р. Могендовича (1957), главная роль кинестезии для кровообращения
заключается в осуществлении моторно-виснеральных рефлексов. При
изучении последних использовали в качестве раздражителя растяжение
проприорецепторов скелетных мышц. При этом имелось в виду, что
растяжение скелетных мышц является более адекватным раздражителем
проприорецепторов по сравнению с сокращением, заключающимся в том, что
оно, якобы, исключает влияние на кровообращение..
Следует
заметить, что скелетная мышца в литературе не рассматривалась как
самостоятельный присасывающе-нагнетательный насос, а принималась в
качестве рабочего органа, перфузируемого кровью. В последнее время
стало известно, что она наряду с этим является насосом,, который, как
сказано выше, функционирует не только при сокращении, но и в
физиологическом покое. В связи с этим представляло интерес изучить
вопрос о том, функционирует ли скелетная мышца и в процессе
-ритмического растяжения как присасывающе-нагнетательный насос;
принимает ли растяжение наряду с сокращением активное участие как в
самообеспечении скелетных мышц, так и в кровообращении организма в
целом или только служит раздражителем проприорецепторов в
осуществлении моторно-висцеральных рефлексов.
ВЛИЯНИЕ ФИЗИЧЕСКОЙ ТРЕНИРОВКИ
НА СЕРДЕЧНО-СОСУДИСТУЮ И МЫШЕЧНУЮ СИСТЕМЫ
В связи с
прогрессирующей заменой в труде и в быту мышечной деятельности
человека работой машин происходит ограничение двигательной активности,
что приводит к гипокинезии организма. Как известно, пониженная
двигательная активность ухудшает состояние не только мышечной системы в
виде атрофических изменений двигательного аппарата (Панов и др., 1969;
Порту-галов и др., 1969), но и сердечно-сосудистой, а также многих
других систем организма.
Физическая
же культура и спорт являются одним из эффективных средств профилактики
и лечения как опорно-двигательного аппарата, так и сердечно-сосудистых
заболеваний. В последнее время при лечении инфаркта
|
|||||
|
24
|
|||||
|
миокарда все
шире применяется не охранительный, постельный режим покоя, а активный
двигательный режим и физические упражнения в виде лечебной гимнастики.
Лечебная
гимнастика, проводившаяся методом интервальных тренировок, вызывала у
больных инфарктом миокарда существенные сдвиги гемодинамики: урежение
пульса, снижение минимального и среднего артериального давлений,
увеличение ударного объема крови и адекватное уменьшение удельного
сопротивления сосудов, что свидетельствует о благоприятном влиянии
лечебной гимнастики на сердечно-сосудистую систему (Танеев, 1977).
Все органы и
системы по структуре и функциям рассчитаны на оптимальную активность,
а не на покой, и могут достигать полного развития лишь в условиях
достаточно интенсивной деятельности. Активная физическая
деятельность, совершаемая в оптимальном объеме и интенсивности в
течение всей жизни человека, способствует повышению экономичности и
надежности сердечнососудистой системы и всего организма в целом
(Чеботарев, Коркушко, Калиновская, 1969).
Применение
максимальных физических нагрузок не всегда может быть полезным. По
некоторым данным интенсивная физическая нагрузка приводит к
неполноценному кровоснабжению миокарда, недостаточному поступлению
нуклеиновых кислот, в мышце наблюдаются резкие сдвиги
ангиоархитектоники, особенно со стороны капилляров (Алашеева, 1975).
Показано,
что ступенчатое увеличение мышечных нагрузок при систематическом их
применении имеет существенное преимущество перед равномерным и
значительно повышает степень адаптации к ней (Пыжова, 1972, 1973), а
эффективность адаптации в свою очередь может изменяться в зависимости
от распределения одной и той же нагрузки в течение дня (Яковлев,
Яковлева, 1971).
А. П.
Сорокин и др. (1973), занимаясь изучением дозировки физической
нагрузки в эксперименте, обратили внимание на то, что время выполнения
нагрузки при одинаковой интенсивности не точно отражает ее величину и
вызывает различные по степени ответные реакции сердечно-сосудистой,
дыхательной и других систем организма. Они экспериментально показали,
что желательно выбирать время действия нагрузки при постоянной
интенсивности и контролировать ее дозировку с учетом ди-
|
|||||
|
25.
|
|||||
|
намики параметров сердечно-сосудистой и дыхательной систем.
Изучению
влияния физической нагрузки на сердечнососудистую систему посвящено
большое количество ра^ бо* (Духовичный, 1964; Озолинь, Порцик, 1970;
Пирого- ' ва, 1970; Сорокин и др., 1973 и др.).
Физическая
тренированность, как считают В. В. Парии, Ф. 3. Меерсон'(1965),
обеспечивает экономное осуществление обменных процессов, быструю и
полную мобилизацию функции кровообращения.
Адаптация
сердца к повторным физическим нагрузкам осуществляется за счет,
во-первых, функционального расширения полостей сердца, во-вторых,
физиологической гипертрофии миокарда в результате увеличения толщины
его мышечных волокон (Reindell et al., 1960; Комадел, Барта, 1968 и
др.).
Под влиянием
тренировки поддержание минутного объема кровообращения может
происходить как за счет увеличения систолического объема, так и
вследствие повышения частоты ритма сердечной деятельности
(Абер-берг-Ауешканпе, 1977).
D. J.
McCloskey (1976) показано, что при динамической физической нагрузке
происходит увеличение минутного объема сердца, ударного объема,
пульса, повышается систолическое давление, т. е. отмечается нагрузка
на сердце. При статической физической нагрузке минутный объем сердца и
пульс повышались незначительно, а системное и диастолическое давления
существенно увеличивались.
Физическая
тренировка вызывает повышение уровня артериального давления в среднем
на 10—20%. Повышение артериального давления при мышечной работе может
объясняться рефлекторными влияниями, возникающими в самой работающей
скелетной мышце. В ре-t зультате. физической
тренировки возрастает функциональная зависимость между локомоторной
системой, с одной стороны, центральной нервной и сердечно-сосудистой
системами с другой (Могендович, 1972).
По данным М.
И. Гуревича, С. А. Бернштейна, Н. В. Ильчевича (1970), при умеренной
физической нагрузке увеличение минутного объема сердца достигалось
преимущественно за счет ритма сердечных сокращений; при повышенной же
— вследствие увеличения ударного объема. Они считают, что сдвиги
гемодинамики достигают
26
|
||||||
|
максимального
уровня в процессе выполнения самой физической работы. Важнейшая роль в
повышении системного артериального давления принадлежит увеличению
минутного объема сердца, изменение которого связано в основном с
возрастанием ударного объема, а не ритма сердечных сокращений.
В состоянии
покоя частота сердечных сокращений у тренированных животных меньше, а
масса сердца больше, чем у нетренированных. Эти изменения зависят как
от возраста животных, так и от интенсивности тренировки (Ракочи,
1971). У хорошо тренированных спортсменов в состоянии покоя обнаружено
снижение минутного объема кровообращения (Кузьмина, 1955; Mellerowicz,
1960).
Под влиянием
адаптации к физическим нагрузкам возрастает максимальная насосная
функция изолированного сердца (Капелько, Меерсон, Шагинова, 1973;
Ка-лелько, Гибер, 1977).
Ф. 3.
Меерсон, В. И. Капелько, К- Пфайффер (1976) обнаружили, что при
физической тренировке происходит увеличение левого желудочка, амплитуды
сокращений папиллярных мышц. Они полагают, что повышение
функциональной возможности сердечной мышцы под влиянием адаптации
обеспечивается увеличением АТФ-азной активности миозина и активации
систем ионного и кислородного транспорта. Ими было показано, что после
регулярной физической тренировки растяжимость миокарда значительно
возрастает и максимальная амплитуда сокращений достигается при меньшей
растягивающей силе.
Систематическая
физическая тренировка наряду с функциональными изменениями в
деятельности сердца лриводит и к морфологической перестройке миокарда,
которая выражается в повышении соотношения числа капилляров к числу
мышечных волокон в сердечной мышце (Bell, Rassmussen, 1974).
Таким
образом, под влиянием физической тренировки в функциях
сердечно-сосудистой системы наблюдаются значительные изменения.
Тренированность оказывает и непосредственное влияние на мышечный
кровоток. Однако данные, полученные при изучении кровотока в мышцах,
весьма противоречивы. Если одни авторы нашли, что физическая работа
вызывает большее усиление мышечного кровотока у малотренированных лиц,
чем у
|
|||||
|
27
|
|||||
|
спортсменов
(Mellerowicz, I960), то, по мнению других, кровообращение интенсивнее в
более тренированных мышцах (Абросимова, 1968). В условиях покоя
кровоток в конечностях у более тренированных спортсменов уменьшается
(Озолинь, Порцик, 1970).
Систематическая
физическая тренировка уменьшает приток крови к неработающим мышцам, что
приводит к более экономному-использованию минутного объема
кровообращения. Наряду с этим установлено, что кровоснабжение
поперечно-полосатой мышечной ткани может меняться в зависимости от вида
физической нагрузки (Petren et al., 1936; Катинас, 1957). При
физической тренировке существенно изменяется тонус скелетных мышц.
Последний в большей степени зависит от длительности и интенсивности
тренировочного процесса. При произвольном напряжении наблюдается
увеличение тонуса мышц, а при произвольном расслаблении — его
уменьшение, благодаря чему увеличивается амплитуда твердости мышц, т.
е. повышаются функциональные возможности мышечной системы (Ефимов,
1947; Кесарева, 1960). Одновременно с функциональными изменениями в кровоснабжении
мышечной ткани после адаптации к физическим нагрузкам происходит и
морфологическая перестройка скелетных мышц, увеличение мышечной массы.
Н. А. Бодрова (1965) обнаружила у молодых щенков, в течение 2—8 мес
тренировавшихся на третбане, изменения в скелетных мышцах конечностей,
которые характеризовались удлинением отдельных мышечных пучков
различного порядка. При этом мелкие и более короткие пучки растут
интенсивнее, чем крупные, а также изменяются группировки волокон в
пучки, что приводит к большому единообразию их по толщине,
увеличению количества соединений между пучками и т. д. Применение
нагрузок до 3-й стадии утомления ведет при неизменной: массе и объеме
мышцы к увеличению толщины мышечных волокон (Денисов, 1973).
После
адаптации к физической нагрузке происходит снижение относительного
количества красных и нарастание белых волокон без изменения количества
промежуточных волокон, особенно в мышцах, на которые приходится
основная двигательная нагрузка. При этом достоверно увеличивается
площадь поперечного сечения промежуточных и белых мышечных волокон,
расположенных в наружных областях мышцы (Дерибас, Филип-
28
|
|||||
|
/
|
|||||
|
ченко,
1974). В функциональном отношении с деятельностью красных волокон
связывают функцию поддержания тонуса, а с деятельностью белых — фазных
сокращений.
Таким
образом, после адаптации к мышечным нагрузкам повышается
функциональная активность мышечных волокон, связанных с фазными
сокращениями.
П. 3. Гудзь
(1968, 1973), изучая морфологическую перестройку скелетных мышц у
животных (белых крыс, собак), обнаружил, что малые физические нагрузки
не вызывают заметных изменений гистоструктуры, а при повышенных
нагрузках происходит гипертрофия и гиперплазия мышечных волокон с их
продольным расщеплением. Перестраивается также и капиллярная сеть
мышцы. Петли капилляров прорастают между вновь образовавшимися
мышечными волокнами. Уже при малых физических нагрузках происходят
увеличение капиллярного русла поперечно-полосатых мышц и возрастание
числа функционирующих капилляров (Brodal, Ingjer, Hermansen, 1977).
Из
приведенных данных литературы следует, что физическая тренировка ведет
к изменениям гистоструктуры скелетных мышц и создает ряд предпосылок
для воспитания, особенно у спортсменов, качеств силы, скорости и
выносливости.
Исходя из
тесной функциональной взаимосвязи и взаимозависимости, существующих
между сердечно-сосудистой и мышечной системами, выяснение роли
скелетных мышц в гемодинамике представляет значительный теоретический
и практический интерес.
Итак, в
организме человека и животных наряду с центральным гемодинамическим
насосом — сердцем — в кровообращении принимают активное участие
многочисленные его помощники — внутримышечные периферические
«сердца». В результате присущей им присасывающей способности они
снижают периферическое сопротивление, а благодаря нагнетательной
функции, проявляющейся при разных видах сокращения, проталкивают кровь
из артерий по внутримышечным капиллярам в вены с силой, превышающей
даже максимальное артериальное давление, увеличивая и ускоряя возврат
_, венозной крови к сердцу, его наполнение, что приводит к
значительному облегчению его работы. В основе микронасосной
деятельности скелетных мышц лежит вибра-
|
|||||
|
29
|
|||||
|
ционный
механизм, заключающийся в том, что асинхронно сокращающиеся мышечные
волокна, воздействуя на внутримышечные сосуды, способствуют продвижению
заключенной в них крови. Описанная выше деятельность внутримышечных
периферических «сердец» изучалась в условиях физиологического покоя,
ритмического, тетани-ческого сокращения мышцы, ее массаже и при
гипотермии, сопровождающейся возникновением холодовой мышечной дрожи.
Но для более
полной характеристики внутримышечных периферических «сердец»
необходимо изучение роли скелетных мышц в гемодинамике организма не
только при различных видах сокращения, но и растяжения. В лаборатории
кровообращения Института физиологии АН БССР исследовалось участие
ритмического растяжения изолированной икроножной мышцы в проявлении
функции внутримышечного периферического «сердца», а также влияние
физической тренировки на эффективность внутримышечного периферического
«сердца» в условиях ритмического растяжения скелетных мышц. Решались
следующие вопросы: 1. Обладает ли скелетная мышца при ритмическом
растяжении нагнетательной функцией и как влияет различная нагрузка (от
1 до 10 кг) на нагнетательный эффект? 2. Если скелетная мышца при
растяжении есть насос, то проявляет ли она присасывающий эффект? 3.
Способна ли скелетная мышца в условиях ритмического растяжения
осуществлять замкнутую циркуляцию крови по искусственному кругу
кровообращения и какова объемная скорость кровотока при этом? 4. Каков
механизм проявления приса-сывающе-нагнетательной функции при
ритмическом растяжении скелетных мышц, изучаемый с помощью
электромиографии, электротономиографии и других фрагментов
исследований? 5. Влияет ли физическая тренировка на
присасывающе-нагнетательную микронасосную способность скелетных мышц
при их растяжении? :
|
||||||||
|
Глава II
|
|||||||
|
МЕТОДИКИ И ОБЪЕКТЫ ИССЛЕДОВАНИЙ
|
|||||||
|
Вся
экспериментальная часть работы основана на проведении острых опытов,
выполненных на изолированной икроножной мышце собак при сохранении
нервных и сосудистых связей с организмом, а также на полностью
изолированных икроножных мышцах животных. Наряду с острыми опытами
проведены хронические исследования на собаках, подвергавшихся
систематической физической тренировке, с целью изучения возможности
повышения эффективности микронасосной способности скелетных мышц в
условиях их растяжения и влияния последних на показатели
сердечно-сосудистой системы у собак в целом.
ИЗОЛИРОВАНИЕ ИКРОНОЖНОЙ МЫШЦЫ
Работа
выполнена на 89 беспородных собаках, находившихся под
морфинно-гексеналовым наркозом. Перед опытом на 30—40 мин собакам
вводился подкожно 2%-ный раствор соляно-кислого морфина из расчета 0,5
мл на 1 кг массы животного, затем в процессе опыта внутрибрюшинно
вводился 5%-ный раствор гексенала. Подготовка животного к исследованию
проводилась следующим образом: животное с помощью лямок укрепляли на
операционном столе в горизонтальном положении на спине. Шерсть с
задней конечности тщательно состригали, и операционное поле
обрабатывали настойкой йода. Скальпелем производили послойный разрез
подкожно-жировой клетчатки по медиальной поверхности. На мышцу
gastrocnemius попарно накладывали жгуты, между которыми осуществляли
полное пересечение тканей. Тупым способом вычленяли икроножную мышцу
из окружающих тканей, перевязывали все приходящие и отходящие от мышцы
сосуды, за исключением бедренных артерии и вены. Бедренные артерия и
вена
|
|||||||
|
31
|
|||||||
|
в условиях
физиологического покоя необходима в качестве контроля при применении
препаратов кураре (ди-плацина). По ее исчезновению можно судить о
начале действия диплацина на нервно-мышечные синапсы скелетных мышц.
Возникающие
в мышце потенциалы действия отводились при помощи игольчатых
электродов, расположенных на расстоянии 1 .см друг от друга, и
подавались на усилитель переменного тока БПУ 4-01. Запись
электро-миограммы осуществлялась с помощью чернильнопи-шушего прибора
УСЧ 8-03 со скоростью движения бумаги 50, 100 мм/с. Количественный
анализ электромиограм-мы проводился визуально по методу, описанному Р.
В. Персон (1969)'.
Данные
электромиограммы в известной степени отражают тонические напряжения
скелетных мышц. В дополнение к этому методу был применен также способ
измерения тонуса икроножной мышцы как в состоянии «видимого» покоя, так
и при различной силе растяжения с помощью электромиотонометра ЭМ-3. Щуп
устанавливали на область икроножной мышцы, а показания шкалы с
помощью графика переводились в г/см2.
Приведенный
комплекс методик использовался для исследования гемодинамики
изолированной икроножной мышцы в условиях ее растяжения и изучения
некоторых вопросов механизма влияния растягивающихся скелетных мышц на
внутриорганный кровоток.
|
||||||
|
Глава III
|
||||||
|
НАГНЕТАТЕЛЬНАЯ ФУНКЦИЯ
ВНУТРИМЫШЕЧНОГО
ПЕРИФЕРИЧЕСКОГО «СЕРДЦА»
В УСЛОВИЯХ РИТМИЧЕСКОГО РАСТЯЖЕНИЯ
ИЗОЛИРОВАННОЙ СКЕЛЕТНОЙ МЫШЦЫ
С помощью
описанных выше методов исследования получены факты, которые дают
возможность в количественных показателях давления и объема
перекачиваемой крови изучать деятельность внутримышечных
периферических «сердец» не только при различных видах сокращения, но и
растяжении.
ДИНАМИКА ЗАСТОЙНОГО ВЕНОЗНОГО ДАВЛЕНИЯ
ПРИ ПРЕГРАЖДЕНИИ ВЕНОЗНОГО ОТТОКА
ИЗ СОСУДОВ ИЗОЛИРОВАННОЙ ИКРОНОЖНОЙ МЫШЦЫ,
НАХОДЯЩЕЙСЯ В УСЛОВИЯХ
ФИЗИОЛОГИЧЕСКОГО ПОКОЯ
Прежде всего
было проведено изучение особенностей гемодинамики в изолированной
икроножной мышце собаки при сохранении нервных и сосудистых связей с
организмом в условиях покоя без отягощения ее грузами.
С этой целью
в острых опытах наблюдали динамику застойного венозного давления при
пережатии бедренной вены икроножной мышцы, находящейся в условиях
физиологического покоя. Эти опыты служили контролем для сопоставления
их с гемодинамикой в условиях растяжения икроножной мышцы.
Как показали
результаты исследований, застойное венозное давление в условиях
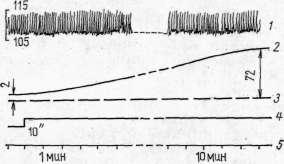
физиологического покоя мышцы медленно повышалось (рис. 3), но, как
правило, не достигало уровня максимального артериального давления.
Законы
гемодинамики, учение о движении крови по сосудистой системе являются
общими с законами гидродинамики— учением о движении жидкости.
Вследствие этого по закону сообщающихся сосудов застойное венозное
давление при преграждении венозного оттока должно было бы достичь
уровня максимального артериально-
|
||||||
|
гг
|
||||||
|
го давления.
Однако периферическое сопротивление в артериолах и капиллярах,
эластические и упругие свойства сосудистой стенки, вязкость крови и
другие особенности крови й сосудов мышцы создают значительное,
сопротивление току крови, на преодоление которого расходуется основное
(85%) количество энергии сердца (Бабский и др., 1966). Поэтому в наших
экспериментах не наблюдалось выравнивания уровней застойного венозного
и максимального артериального давлений (табл. 1).
|
||||||||||||
 |
||||||||||||
|
Рис. 3. Динамика застойного венозного давления в условиях физиологического покоя изолированной икроножной мышцы собаки: 1 — кимограмма артериального давления; 2 — кимограмма застойного венозного давления, 3 — нулевая линия, 4 — отметка наложения сосудистого зажима на бедренную вену, 5 — отметка времени
|
||||||||||||
|
С другой
стороны, для выравнивания уровней застойного венозного и максимального
артериального давлений большую роль играет фактор времени. Возможно,
что при более продолжительном преграждении венозного оттока по
сравнению со временем, применяемым в наших экспериментах (10 мин), и
произошло бы; вырав^. нивание. Но мы имеем дело с живым организмом, а,
как известно, длительный венозный застой крови ведет к нарушению
обмена веществ в тканях, что в свою очередь вызывает отек тканей, их
набухание, в том числе и набухание стенок кровеносных сосудов, которое
приводит к сужению их просвета. Такое состояние не является
физиологическим, и поэтому длительное преграждение венозного оттока
из мышцы не может использоваться для решения поставленных перед данной
работой задач.
Как следует
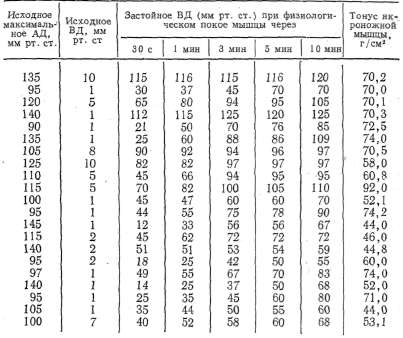
из табл. 1, максимальное артериальное давление (АД) у собак разного
возраста в среднем было 114+3,177 мм рт.ст., венозное давление (ВД)
перед
за
|
||||||||||||
|
Таблица 1 Данамика застойного венозного давления при преграждении венозного оттока крови из сосудов икроножной мышцы, находящейся в условиях физиологического покоя
|
||||||||
 |
||||||||
|
опытом
равнялось 3,2+0,519 мм рт. ст. При преграждении венозного оттока
застойное венозное давление неуклонно повышалось и к 10-й минуте
достигало 84,4+ +4,043 мм рт. ст., т. е. не превышало максимального
артериального давления. Причем в одних опытах (3, 4, 10) застойное
венозное давление повышалось довольно быстро и приближалось по своему
значению к максимальному артериальному давлению, а в других (13, 14,
15, 18) повышалось очень медленно и к концу эксперимента было
значительно ниже максимального артериального давления.
Таким
образом, диапазон повышения застойного венозного давления мышцы,
находящейся в условиях физиологического покоя, был довольно
значительным и колебался у разных животных от 55 мм рт. ст. до 120—
125 мм рт. ст. (см. табл. 1).
По всей вероятности, такой большой диапазон повы-
|
||||||||
|
39
|
||||||||
|
(пения
застойного венозного давления .у ..собак обусловлен разным
функциональным состоянием скелетных мышц, степенью их тренированности,
неодинаковым тонусом, который объясняется большинством авторов
(Юсевич, 1963; Махмудов, 1965; Водолазский, 1966; Жуков, 1969 и др.)
как проявление активного состояния мышцы, разным исходным уровнем
артериального давления и рядом других причин.
У собак с
хорошо выраженным тонусом скелетной мышцы (см. табл. 1) (6, 10, 12, 17)
застойное венозное давление повышалось более стремительно и достигало
больших величин, чем у собак-со слабо выраженным мышечным тонусом.
Дальнейшие
исследования были продолжены на собаках, у которых изучались изменения
застойного венозного давления под влиянием растяжения икроножных мышц
путем отягощения их различными грузами.
ПРЕВЫШЕНИЕ ЗАСТОЙНЫМ ВЕНОЗНЫМ ДАВЛЕНИЕМ МАКСИМАЛЬНОГО АРТЕРИАЛЬНОГО ПРИ РИТМИЧЕСКОМ РАСТЯЖЕНИИ ИЗОЛИРОВАННОЙ СКЕЛЕТНОЙ МЫШЦЫ
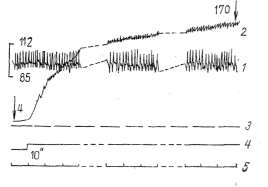
Для
выяснения специфики гемодинамики в икроножной мышце, подвергавшейся
ритмическому растяжению с частотой 1 растяжение в секунду, был взят
груз 6 кг.
Изолированная
икроножная мышца при сохранении нервных и сосудистых связей с
организмом при ее растяжении проявила себя как мощный нагнетательный
насос. В данных условиях работы мышцы застойное венозное давление
росло более стремительно, чем при преграждении венозного оттока из
скелетной мышцы в условиях ее физиологического покоя. Оно не только
достигало максимального артериального давления, но и способно было
превышать его на значительную величину (рис. 4). Как видно из табл. 2,
максимальное артериальное давление в этом, случае составляло
106,8+3,104 мм рт. ст., а застойное венозное увеличивалось до
158,6+4,222 мм рт. ст. с превышением максимального артериального
давления на 52,2 мм рт. ст., т.е. на 48,9%.
Это
свидетельствует о том, что не только при различных видах сокращения,
массаже, но и при ритмическом растяжении скелетная мышца работает как
эффективный
40
|
||||||
|
нагнетательный
насос. Этот факт не является какой-то редкой случайностью. Он
наблюдался во всех без исключения проводившихся экспериментах, где
растяжение икроножной мышцы осуществлялось грузом 6 кг (см. табл. 2).
При уровне
максимального артериального давления 106,8+3,104 мм рт. ст. и исходном
венозном давлении 4,6+1,118 мм рт. ст. застойное венозное давление уже
через 30 С после наложения, сосудистого зажима на бед-
|
|||||||||||
 |
|||||||||||
|
1мин Змии 5мцн
Рис. 4.
Динамика застойного венозного давления при ритмическом растяжении
икроножной мышцы грузом 6 кг: ) — кодограмма артериального давления, 2 — кимограмма застойного венозного давления, 3 — нулевая линия, 4 — отметка наложения сосудистого зажима на бедренную вену, 5 — отметка времени
|
|||||||||||
|
ренную вену
достигало максимального артериального уровня, к 1-й минуте превысило
его на 23 мм рт. ст., к 3-й — на 40, а к концу 5-й минуты — на 52 мм
рт. ст. Таким образом, уже через 3 мин застойное венозное давление
достоверно (Р<0,001) превышало уровень максимального артериального.
Следовательно,
не только при сокращении, но и растяжении, при котором сокращение
исключалось, проявляется нагнетательная способность скелетной мышцы.
Образно говоря, в работе скелетных мышц как микронасосов нет
«холостого хода». Нагнетательная функция осуществляется и в фазу
сокращения и в фазу растяжения.
Как в
условиях физиологического покоя, так и в исследованиях с применением
растяжения скелетных мышц обнаруживаются индивидуальные различия в эф-
|
|||||||||||
|
41
|
|||||||||||
|
Таблиц3 2
|
|||||||||
|
Динамика застойного венозного давления при ритмическом
растяжении икроножной мышцы грузом 6 кг й частотой 1
растяжение в секунду, мм рт. ст.
|
|||||||||
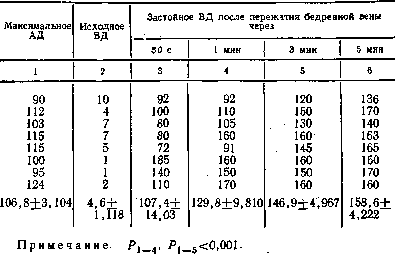 |
|||||||||
|
фективности
деятельности икроножных мышц как микронасосов. Так, более дряблые, с
меньшим тонусом икроножные мышцы (3, 8) в процессе своей деятельности
при растяжении вызывали превышение застойным венозным давлением уровня
максимального артериального на 36—37 мм рт. ст., в то время как другие,
более плотные, с высоким исходным тонусом (6, 7) — на 65 мм рт. ст.,
т.е. в два раза больше. Эти данные еще раз свидетельствуют о
зависимости насосной деятельности скелетных мышц от их исходного
функционального состояния и, вероятно, тренированности, что в свою
очередь говорит о возможности внешнего воздействия на насосную
способность скелетных мышц и управления их деятельностью.
Наряду с
этими наблюдениями возник вопрос о том, как-влияет различная сила
растяжения скелетных мышц на их насосную эффективность, так как .взятый
первоначально груз в 6 кг для растяжения скелетных мышц способствовал
лишь выявлению самого феномена — насосной деятельности в этих
условиях. Представляло интерес выяснить, будет ли проявляться насосная
спо-
42
|
|||||||||
|
собность
скелетных мышц при растяжении различными грузами и какова ее
эффективность. - Для выяснения этих вопросов были проведены дальнейшие
исследования.
ЗАВИСИМОСТЬ ВЕЛИЧИНЫ ЗАСТОЙНОГО ВЕНОЗНОГО ДАВЛЕНИЯ ОТ СИЛЫ РИТМИЧЕСКОГО РАСТЯЖЕНИЯ ИЗОЛИРОВАННОЙ ИКРОНОЖНОЙ МЫШЦЫ
Изучение
закономерности повышения уровня застойного венозного давления от
нагрузки проводилось в диапазоне 1 —10 кг с частотой 1 растяжение в
секунду.
Обнаружено,
что ритмическое растяжение мышцы грузом 1—2 кг повышало застойное
венозное давление, однако оно не достигало уровня максимального
артериального (табл. 3). Динамика повышения застойного венозного
давления при растяжении указанным грузом близка к характеру повышения
такового в условиях физиологического покоя.
При
растяжении икроножной мышцы нагрузкой 3, 4 кг застойное венозное
давление повышалось более стремительно и к концу 5-й минуты начинало,
хотя и незначительно, но все же превышать уровень максимального
артериального давления (табл. 4, 5).
Таблица 3
|
|||||||||
|
Застойное венозное давление при ритмическом растяжении
икроножной мышцы грузом 1—2 кг и частотой 1 растяжение
в секунду, мм рт. ст.
|
|||||||||
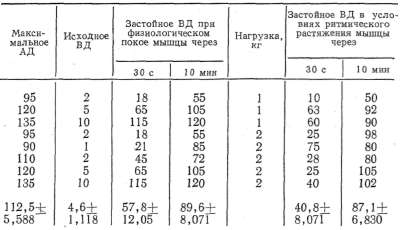 |
|||||||||
|
43
|
|||||||||
|
Таблица 4
|
||||||||||||
|
Застойное венозное давление при ритмическом растяжении
икроножной мышцы грузом 3 кг и частотой 1 растяжение
в секунду, мм рт. ст.
|
||||||||||||
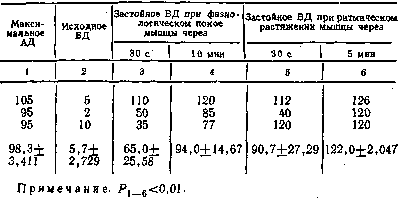 |
||||||||||||
|
Таблица 5
|
||||||||||||
|
Застойное венозное давление крови при ритмическом растяжении мышцы грузом 4 кг И частотой I растяжение в секунду, мм рт. ст.
|
||||||||||||
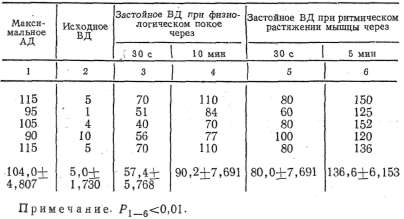 |
||||||||||||
|
Сопоставляя
уровни застойного венозного давления при растяжении грузами 1 и 2 кг,
мы не обнаружили между ними различий, поэтому полученные данные,
сведены в одну таблицу (см. табл. 3). Вероятно, растяжение порядка 1—2
кг является под пороговой величиной. Пороговая величина обнаруживается
в условиях растяжения мышцы грузом 3 кг, когда застойное венозное
|
||||||||||||
|
44
|
||||||||||||
|
Таблица
Застойное венозное давление при ритмическом растяжении
икроножной мышцы грузом 7 кг и частотой 1 растяжение
в секунду, мм рт, ст.
|
|||||||||||||
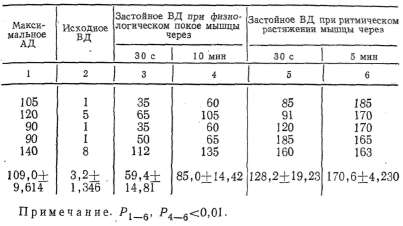 |
|||||||||||||
|
давление
достоверно выше такового при 1—2 кг (см. табл. 4). Растяжение
икроножной мышцы, грузом 4 кг приводило к еще большему превышению
застойным венозным давлением максимального артериального (см. табл. 5).
Дальнейшее увеличение нагрузки до 6 кг давало, как рассмотрено ранее (см. табл. 2), достоверное превыше-
|
|||||||||||||
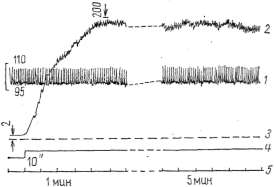 |
|||||||||||||
|
Рис. 5.
Повышение застойного венозного давления при ритмическом растяжении
икроножной мышцы грузом 10 кг: / — артериальное давление, 2 — застойное венозное давление, 3 — нулевая линия, 4 — отметка наложения сосудистого зажима на бедренную вену, 5 —
отметка времени
|
|||||||||||||
|
45
|
|||||||||||||
|
Таблица 7
|
||||||||||||||
|
Динамика застойного венозного давления при ритмическом растяжений
икроножной мышцы грузом 10 кг и частотой 1 растяжение
в секунду, мм рт. ст
|
||||||||||||||
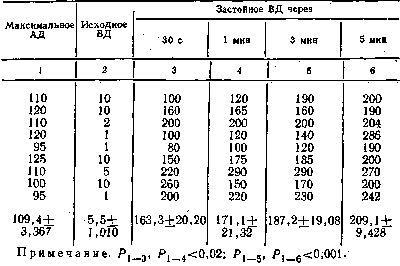 |
||||||||||||||
|
ние
застойным венозным давлением максимального артериального. Оно было еще
больше при 7 кг растяжения икроножной мышцы (табл. 6). А растяжение
скелетных мышц грузом .10 кг приводило к стремительному нарастанию
застойного венозного давления, которое уже через 30 с от начала
растяжения достоверно (Р<0,01) превышало уровень максимального
артериального давления.
|
||||||||||||||
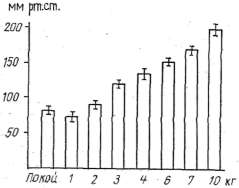 |
||||||||||||||
|
Рис. 6. Зависимость повышения застойного венозного давления от
Силы растяжения
46
|
||||||||||||||
|
I
|
||||||||||||||
|
К концу 1-й
минуты растяжения оно было достоверно выше по сравнению с его значением
в состоянии физиологического покоя мышцы через, 10 мин венозной
окклюзии (рис. 5, табл. 7).
Итак, предел
повышения застойного венозного давления икроножной мышцы, находящейся
под воздействием ритмического растяжения нагрузками в диапазоне от 1
до 10 кг, зависит от силы растяжения. Эта зависи
|
||||||||||||
|
мость
представлена на рис. 6. Возможно, что при дальнейшем увеличении силы
застойное венозное давление будет продолжать возрастать, однако наши
исследования проводились
|
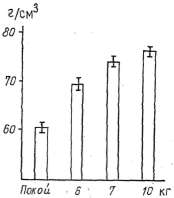 |
|||||||||||
|
Рис. 7. Динамика изменения тонуса икроножной мышцы в условиях ее ритмического растяжения различной силой
|
||||||||||||
|
лишь в
данном диапазоне. Изучение динамики изменения застойного венозного
давления при больших силах растяжения является предметом дальнейшего
специального изучения.
Полученные
результаты свидетельствуют о том, что в условиях данных экспериментов
при ритмическом растяжении икроножной мышцы собаки грузом 1—2 кг
насосную деятельность скелетных мышц обнаружить не удалось.
По-видимому, при незначительном растяжении в работе принимает участие
незначительное количество двигательных единиц, поэтому в мышце
недостаточно реализуется заключенный в ней микронасосный механизм, т.
е. в данном случае создаются условия, близкие к состоянию гипокинезии,
губительной для организма (Коваленко, Гуровский, 1980; Коваленко
(ред.), 1983). Начиная с 3 и до 10 кг нагрузки, скелетная мышца (см.
рис. 6) проявляет явно выраженный нагнетательный эффект, заключающийся
в активном однонаправленной перекачивании крови через обширную
внутримышечную сосудистую сеть.
|
||||||||||||
|
47
|
||||||||||||
|
Зависимость
насосного эффекта от нагрузки, по всей вероятности, объясняется тем,
что при увеличении таковой в работу включается все большее число
одновременно действующих нейромоторных единиц (Кирзон, 1959; Бурсиан,
1972 и др.). в связи с чем усиливается внешнее давление на капилляры и
другие мелкие внутримышечные сосуды. Об усилении внешнего давления на
капилляры могут свидетельствовать и результаты измерения тонуса
изолированной икроножной мышцы, находящейся под влиянием ритмического
растяжения (рис. 7), которые хорошо согласуются с данными литературы
(Стома, 1956, 1958; Степанов, Дементьев, 1967 и др.).
Механизм
нагнетательного эффекта в условиях ритмического растяжения мышцы
довольно сложный. Влияние мышечных волокон на сосуды в этих условиях
определяется многими факторами, среди которых можно выделить скорость
растяжения, функциональное состояние мышцы, ее тип (преобладание в
мышце быстрых или медленных мышечных волокон), силу растяжения и ряд
других.
Таким
образом, изложенными в этой главе исследованиями показано, что
скелетная мышца и при ритмическом- растяжении, осуществляемом с
достаточно сверхпороговой силой, работает как мощный нагнетательный
насос. При каждом растяжении она проталкивает кровь по внутримышечным
сосудам и нагнетает ее на выходе в вену с силой, превышающей
максимальное артериальное давление.
|
||||
|
Глава IV
|
||||||||
|
ПРИСАСЫВАЮЩАЯ ФУНКЦИЯ
ВНУТРИМЫШЕЧНОГО
ПЕРИФЕРИЧЕСКОГО «СЕРДЦА»
В УСЛОВИЯХ РИТМИЧЕСКОГО РАСТЯЖЕНИЯ
ИЗОЛИРОВАННОЙ СКЕЛЕТНОЙ МЫШЦЫ
В предыдущей
главе описан суммарный нагнетательный эффект изучаемого
внутримышечного периферического «сердца», осуществлявшийся с учетом
деятельности центрального насоса — сердца, т. е. сил vis a tergo. При
преграждении венозного оттока застойное венозное давление повышалось в
условиях поступления под высоким давлением артериальной крови,
.нагнетаемой сердцем. И лишь факт превышения застойным венозным
давлением уровня максимального артериального давал полное и несомненно
объективное основание говорить о том, что на пути от сердца находится
дополнительный самостоятельный насос. Он способен значительно
прибавлять давление венозной крови к тому, которое получает от сердца.
Для
исключения сил vis a tergo в условиях острого опыта на изолированной
икроножной мышце собак проведена серия экспериментов с одновременным
преграждением как артериального притока, так и венозного оттока крови
из сосудов икроножной мышцы, находящейся в условиях физиологического
покоя и при ее ритмическом растяжении грузами 7, 10 кг.
Как показали
результаты исследований, наблюдались два варианта динамики изменения
артериального и венозного давлений при физиологическом покое мышцы. В
первом случае артериальное давление резко снижалось, а венозное
оставалось на исходном уровне или повышалось на несколько мм рт. ст.
(рис. 8, а); во втором случае артериальное давление падало, а венозное повышалось так, что образовывался небольшой перекрест (рис. 8, б). Такая
вариабельность в экспериментах с преграждением как артериального
притока, так и венозного оттока крови из икроножной мышцы объясняется
разным исходным тонусом мышц.
При ритмическом растяжении икроножной мышцы
|
||||||||
|
i. Зак. 837
|
||||||||
|
49
|
||||||||
|
грузом 7 кг
артериальное давление значительно понижалось и через 5 мин равнялось
15—20 мм рт.ст., а венозное повышалось, достигая 70—80 мм рт.ст.,
превышая тем самым давление крови, содержащейся в артерии (табл. 8).
Одновременное
преграждение как артериального притока крови в мышце, так и венозного
оттока крови из ее сосудов при ритмическом растяжении грузом 10 кг
приводило к стремительному повышению застойного ве-
|
|||||||||
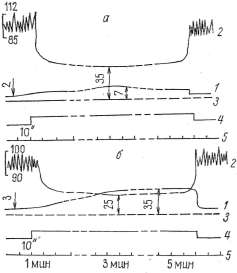 |
|||||||||
|
Рис. 8. Изменения застойного венозного и максимального артериального давления при физиологическом покое икроножной мышцы: 1 — застойное венозное давление, 2 — артериальное давление, 3 — нулевая линия, 4 — отметка наложения и снятия сосудистых зажимов с бедренных артерии и вены, 5 — отметка времени
нозного
давления, которое уже через 30 с от начала эксперимента достигало
120—210 мм рт. ст., и падению давления в бедренной артерии вплоть до
отрицательных величин (рис. 9, см. табл. 8).
Из табл. 8
следует, что и в этих условиях эксперимента как нагнетательная, так и
присасывающая функция икроножных мышц собак различна. Одни мышцы (7, 2, 3, 6) обладали присасывающей способностью, у других она была равна 0, а у третьих (7, 8, 9, 10) присасывающий эффект не проявлялся. Нагнетательная функция также была различной, но во всех случаях застойное
|
|||||||||
|
50
|
|||||||||
|
Таблица 8
|
||||||||||||
|
Динамика артериального и венозного давлений при одновременном
преграждении притока и оттока крови из ритмически растягиваемой
икроножной мышцы собак, мм рт, ст.
|
||||||||||||
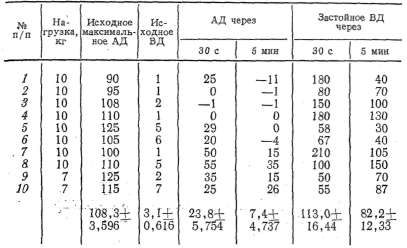 |
||||||||||||
|
венозное
давление превышало давление крови в артериальном сосуде, отключенном
от общего круга кровообращения, а у некоторых животных оно было больше
максимального артериального давления (4, 8). Все это
свидетельствует о значительных индивидуальных осо-' бенностях
проявления присасывающе-нагнетательнои микронасосной способности
скелетными мышцами. "
Таким
образом, в данной серии экспериментов полу-: чены результаты,
однонаправленные с данными предыдущей серии и свидетельствующие о
наличии нагнетательного эффекта, который возрастал при увеличении силы
растяжения. Вместе с тем эти данные позволяют говорить о том, что
наряду с нагнетательным эффектом скелетная мышца при растяжении
обладает и присасывающей функцией, особенно хорошо проявляющейся при
нагрузке 10 кг, когда артериальное давление понижалось до —11 мм рт.
ст. (см. рис. 9).
Общеизвестно,
что основную роль в продвижении крови по большому и малому кругу
кровообращения выполняет сердце, которое является основным
гемодина-мическим насосом, обладающим большой нагнетательной функцией,
активной систолой, но пассивной диастолой,
|
||||||||||||
|
5 Г
|
||||||||||||
|
4'
|
||||||||||||
|
поскольку
окончательно не выяснено, проявляет ли сердце присасывающую
способность. А вот скелетная мышца, как удалось экспериментально
показать, при ритмическом растяжении обладает не только
нагнетательным, но и присасывающим эффектом, активно проталкивает кровь
к сердцу, что делает ее по основным признакам участия в гемодинамике
периферическим «сердцем».
Следовательно, скелетная мышца и в процессе ритмического растяжения является гемодинамическим при-
|
|||||||||||
 |
|||||||||||
|
1MUH 3MUH - 5MUH
Рис. 9.
Кимограмма изменений застойного венозного и максимального
артериального давлений при ритмическом растяжении икроножной мышцы
грузом 10 кг: 1— артериальное давление, 2 — застойное венозное давление, 3 — нулевая линия, 4 — отметка наложения и снятия сосудистых зажимов на бедренные артерию и вену, 5 — отметка
времени
|
|||||||||||
|
сасывающе-нагнетательным
насосом, действующим в помощь сердцу, особенно при выполнении
интенсивной мышечной работы, требующей значительных мышечных усилий.
Таким образом, ритмическое растяжение, являющееся пассивным элементом
двигательных актов, активно участвует в кровообращении организма.
Скелетная
мышца не только при сокращении, но и ритмическом растяжении выполняет
функцию гемодина-мического насоса. Она не остается простым
перфузируе-мым органом, как все другие органы тела человека, а в
современном понимании представляет собой самостоятельный
присасывающе-нагнетательный насос, орган, самообеспечивающийся и
помогающий сердцу.
На основании
проведенных исследований выявлен факт существования
нагнетательно-присасывающей способности скелетных мышц в условиях их
растяжения. Однако эффективность микронасосной деятельности изучалась
по показателям давления. Представлялось инте-
|
|||||||||||
|
52
|
|||||||||||
|
ресным
выяснить насосную эффективность скелетной мышцы при различной силе
растяжения по количеству проталкиваемой полностью изолированной
икроножной мышцей крови, т. е. по объему крови, проходящей через мышцу
в единицу времени.
Изучение
этого вопроса необходимо в связи с тем, что деятельность периферических
«сердец» направлена не только на создание градиента давления крови, но
и на присасывание и нагнетание надлежащего ее количества, необходимого
как для использования самой мышцей, так и для доставки ее по
назначению, т. е. для возврата венозной крови к сердцу с целью его
наполнения.
|
||||
|
Глава V
|
||||||
|
ЭФФЕКТИВНОСТЬ ОБЪЕМНОЙ
НАСОСНОЙ ФУНКЦИИ ВНУТРИМЫШЕЧНОГО
ПЕРИФЕРИЧЕСКОГО «СЕРДЦА»
ПРИ РИТМИЧЕСКОМ РАСТЯЖЕНИИ
ИЗОЛИРОВАННОЙ СКЕЛЕТНОЙ МЫШЦЫ
Обнаруженная
присасывающе-нагнетательная функция скелетных мышц при их ритмическом
растяжении различными грузами весьма эффективна по показателям
отрицательного (присасывающего) и положительного (нагнетательного)
давлений. Последнее превышает даже исходное артериальное давление.
Эффективность
деятельности внутримышечного периферического «сердца» при растяжении
можно выражать не только в показателях присасывающего артериального и
нагнетательного застойного венозного давлений (мм рт. ст.), но и в
единицах объема перекачиваемой крови (мл), что характеризует величину
объемной скорости кровотока через внутримышечное периферическое
«сердце» при растяжении скелетных мышц.
Для этого
икроножную мышцу полностью изолировали и подключали к искусственно
созданному замкнутому кругу кровообращения (см. рис. 2). Как только
икроножную мышцу отключали от большого круга кровообращения и
подключали к искусственному кругу с одновременным ее растяжением
различными грузами — 6, 8, 10 кг и частотой 1 растяжение в секунду,
кровь, содержащаяся в сосудах мышцы, устремлялась через венозную
канюлю по искусственному кругу кровообращения в сторону артерии. В
момент, когда кровь проходила через мерную трубку емкостью 0,5 см3,
отмечали время, за которое мышца прогоняла кровь по этой трубке. По
формуле (см. гл. II) высчитывали объемную скорость кровотока.
В результате
обнаружено, что скелетная мышца и по показателям объема перекачиваемой
крови в процессе ритмического растяжения функционирует как
самостоятельное периферическое «сердце». Ритмически растягиваемая
скелетная мышца в процессе своей деятельности, воздействуя на
капилляры, продвигает кровь из бедрен-
|
||||||
|
54
|
||||||
|
ной вены по
искусственному кругу в бедренную артерию. Таким образом, мышца,
отключенная от нагнетательной системы — сердца, не только при
сокращении, но и при растяжении сама себя снабжает кровью. Это еще раз
подтверждает факт о том, что растяжение скелетной мышцы есть активный
процесс, способствующий продвижению крови и помогающий работе сердца.
Доказано,
что интенсивность кровотока в мышцах зависит от морфологической
структуры мышечной ткани, от функциональных и биохимических
особенностей их волокон. Известно, что особенностями метаболизма
волокон двух типов определяется число кровеносных сосудов в них, а
следовательно, и интенсивность кровотока как в покое, так и при
мышечных сокращениях. Установлено, что в красных мышцах кровоток в
покое сильнее, чем в белых (Folkov, Halicka, 1968; Hilton et al.,
1970). Помимо этого, объемная скорость кровотока, зависит от
упруговязких свойств сосудистой стенки (тонус, эластичность, величина
просвета), количества циркулирующей крови, физических и химических
свойств крови и др.
По данным Б.
Фолкова, Е. Нила, кровоток в скелетных мышцах в состоянии покоя
составляет около 20% , минутного объема кровообращения, а при тяжелой
физической работе может повышаться до 88%. Объем протекающей через
мышцу крови увеличивается с 0,02— 0,06% от общего объема мышцы в покое
до 5,5—15% при работе. При интенсивной мышечной деятельности кровоток в
них может увеличиваться в 12—17 раз по сравнению с покоем (Barkroft,
Swan, 1953; Amery, Bos-saert, Werstraete, 1969).
P. Л. Титиевская (1964) показала, что объемная скорость кровотока предплечья человека в покое равна 2,84+0,19 мл/мин-100 см3
ткани. В. И. Тхоревский (1968), измеряя объемную скорость кровотока
предплечья методом венозной окклюзионной плетизмографии, нашел, что в
покое она составляет 4,5±0,23 мл/мин-100 см3 ткани. Считают,
что в расчете на 100 г мышц предплечья в покое протекает в минуту
1,8—9,6 мл крови, а при работе количество крови увеличивается более чем
на 100 мл.
По некоторым литературным данным, при динамической работе объемная скорость кровотока в мышце равна 20—50 мл/мин-100 см3 ткани (Black, 1959), а при статической (с силой 10—70% от максимальной произволь-
55
|
||||
|
ной) кровоток в предплечье составляет 10—40 мл/мин X Х100 см3 ткани (Humphreys, Lind, 1963; Озолинь, Пор-цик, 1970).
Однако
эти,данные не могут быть использованы для сравнения с результатами,
полученными нами, поскольку интенсивность кровотока как в покое, так и
при мышечной работе измерялась при сохранении связи с основным
насосом — сердцем, постоянно нагнетающим кровь в мышцы, а последние
рассматривались просто как пер-фузируемые органы, рабочая гиперемия в
которых развивается в силу падения тонуса и расширения сосудов. В
наших исследованиях измерялась объемная скорость кровотока в мышце,
полностью изолированной от большого круга кровообращения, т. е.
измерялась та скорость тока крови, которую она развивает сама в
процессе своей деятельности, без какой-либо посторонней помощи. При
этом сокращение исключалось, а объемная скорость кровотока изучалась
только в условиях ритмического растяжения. В этих условиях она
оказалась зависимой от нагрузки: при грузе 6 кг она равнялась 2,2
мл/мин-100 см3 ткани, при нагрузке 8 кг — 2,96, при грузе 10 — 3,25 мл/мин-100 см3 ткани.
Ранее
проведенными исследованиями (Аринчин, Нед-вецкая, Кабуш, 1975)
показано, что полностью изолированная икроножная мышца, замкнутая на
искусственный круг кровообращения, способна самостоятельно
обеспечивать однонаправленную циркуляцию крови при ритмических, а
также тетанических сокращениях. Объемная скорость кровотока при
ритмических сокращениях была 1,94±0,368 мл/мин-100 см3, а при тетанических меньше— 0,637+0,193 мл/мин-100 см3
ткани. Следовательно, эффективность насосной функции скелетных мышц при
ритмическом растяжении оказалась значительно выше, чем при различного
рода сокращениях.
Данные
исследования еще раз подтверждают существование активной микронасосной
функции скелетных мышц, которая проявляется не только в виде
развиваемого им высокого венозного давления, превышающего максимальное
артериальное, но и в продвижении определенного объема кро'ви по
искусственному замкнутому кругу кровообращения.
|
||||
|
Глава VI
|
||||||
|
К ВОПРОСУ О МЕХАНИЗМЕ
ПРИСАСЫВАЮЩЕ-НАГНЕТАЛЬНОЙ
ДЕЯТЕЛЬНОСТИ ВНУТРИМЫШЕЧНЫХ
ПЕРИФЕРИЧЕСКИХ «СЕРДЕЦ»
В УСЛОВИЯХ РИТМИЧЕСКОГО РАСТЯЖЕНИЯ
ИЗОЛИРОВАННЫХ СКЕЛЕТНЫХ МЫШЦ
Механизм
функционирования внутримышечных периферических «сердец» довольно
сложен и многогранен. Одним из представлений, в некоторой степени
объясняющим его, является вибрационная гипотеза (Аринчин, 1961 —1978).
Она основана на уже известном факте об асинхронном сокращении мышечных
волокон в условиях физиологического покоя (состояние тонуса), а
особенно во время ритмических и тетанических сокращений. Согласно этой
гипотезе, микронасосная функция осуществляется скелетными мышцами в
результате асинхронных вибрационных воздействий мышечных волокон на
капилляры с образованием механических движений различного типа: в
виде бегущих волн, за счет которых кровь нагнетается из капилляров в
венозную систему; присасывающих, доящих движений и ряда других
факторов. Основные положения вибрационной гипотезы изучались и нашли
подтверждение в условиях сокращения и покоя скелетных мышц (Ермолова,
Наймитенко, 1970; Наймитенко, 1971; Аринчин, Ермолова, Наймитенко,
Недвецкая, 1973; Аринчин, Володько, 1979 и др.). Возникает вопрос о
том, можно ли объяснить с ее позиций механизм микронасосной функции
скелетных мышц при их ритмическом растяжении?
О МЕХАНИЗМЕ ПРИСАСЫВАЮЩЕ-НАГНЕТАТЕЛЬНОЙ ФУНКЦИИ СКЕЛЕТНЫХ МЫШЦ ПРИ РИТМИЧЕСКОМ РАСТЯЖЕНИИ В УСЛОВИЯХ ИХ ДЕНЕРВАЦИИ И КУРАРИЗАЦИИ
Для
выяснения некоторых вопросов, касающихся механизма функционирования
внутримышечных периферических «сердец», поставлены эксперименты на
изолированной икроножной мышце после ее денервации и ку-раризации.
|
||||||
|
57
|
||||||
|
Опыты с
перерезкой седалищного нерва проводились для того, чтобы изучить, какое
влияние оказывает центральная нервная система на микронасосное
свойство скелетных мышц. Оно оценивалось с помощью метода
преграждения венозного оттока крови из ритмически растягиваемой
икроножной мышцы. Известно, что при растяжении мышцы благодаря
осуществлению моторно-висцеральных рефлексов происходит повышение
артериального давления (Квасова, Некрасов, 1951; Колычев, 1959;
Георгиев, 1961 и др.), а вероятно, и рефлекторное сужение сосудов в
самой скелетной мышце, что может оказывать непосредственное влияние на
величину застойного венозного давления. Эффект повышения
артериального давления исчезает после перерезки задних корешков
спинного мозга или денервации мышцы (Квасова, Некрасов, 1951).
Из данных
литературы следует также, что растяжение, воздействуя на
проприорецепторы, оказывает влияние и непосредственно на тонус мышцы,
повышая его (Уфлянд, 1956; Гранит, 1957; Степанов, Дементьева, 1967),
который сам по себе может явиться дополнительным фактором более
значительного увеличения застойного давления.
В связи с
этим применялась перерезка седалищного нерва с целью исключения
рефлекторных влияний как на сосуды мышцы, от изменения которых может
повышаться общее кровяное давление, так и на тонус мышцы, т. е. по
возможности исключались все рефлекторные влияния и оставался лишь
механический компонент — ритмическое растяжение.
Как следует
из табл. 9, застойное венозное давление после денервации икроножной
мышцы собаки и при ритмическом ее растяжении грузом 10 кг повышалось
так же стремительно, достоверно превышая уровень максимального
артериального давления (Р<0,001), как и с сохраненной иннервацией.
Как известно, препараты кураре блокируют нервно-мышечную передачу и тем самым вызывают падение мышечного тонуса (Иванов, 1965).
При
проведении экспериментов с преграждением оттока крови из ритмически
растягиваемой кураризиро-ванной мышцы действие препаратов кураре
контролировалось с помощью регистрации электромиограммы, которая в
состоянии покоя выражалась в виде редких
|
|||||
|
58
|
|||||
|
Та блица 9
Повышение застойного венозного давления при ритмическом растяжении грузом 10 кг денервированной икроножной мышцы
|
||||||||||||
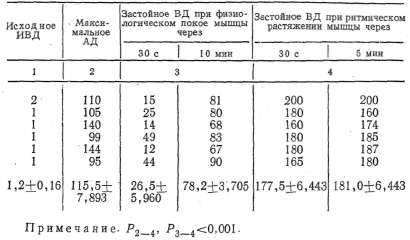 |
||||||||||||
|
осцилляции (18—20 кол/с) с небольшой амплитудой (61,9 мкв) (рис. 10, с).
После введения кураре электромиограмма стала еще более сниженной по частоте амплитуды (рис. 10, б).
При
растяжении икроножной мышцы грузом 3 кг после ее кураризации застойное
венозное давление по сравнению с контрольными исследованиями
существенно снизилось (табл. 10). Опыты на кураризированной мышце
явились дополнительным доказательством в пользу участия активного
механического фактора в микронасосной деятельности скелетных мышц.
Следовательно, нагнетательный эффект в данных вариантах исследований обусловлен не столько рефлек-
|
||||||||||||
 |
||||||||||||
|
Рис. 10. Электромиограмма покоящейся икроножной мышцы собаки: а — ЭМГ до введения диплацина, б — ЭМГ через 20 мин после введения диплацина
|
||||||||||||
|
59
|
||||||||||||
|
торным
влиянием центральной нервной системы на просвет сосудов и тонус мышц,
сколько механическим воз-действие'м растягивающихся мышечных волокон на
внутримышечные сосуды.
Это
заключение еще более подтверждается тем, что объемная скорость
кровотока при ритмическом растяжении грузом 10 кг после денервации
икроножной мышцы собаки была даже большей по сравнению с объемной
скоростью кровотока иннервированной мышцы.
Таким
образом, можно предположить, что при ритмическом растяжении (как и при
сокращении) механизм насосной функции имеет вибрационную природу, т. е.
при растяжении мышечные волокна, асинхронно растягиваясь, оказывают
внешнее давление на капилляры и другие мелкие сосуды и проталкивают
кровь в сторону сердца. При увеличении силы растяжения, с одной
стороны, растет число одновременно растягивающихся единиц, а с
другой, увеличивается давление мышечных волокон на сосуды.
Предпосылкой для такого предположения может служить то, что, как известно,
сокращение (контракция) мышцы состоит из асинхронных сокращений
отдельных мышечных волокон, которые входят в различные моторные
единицы, неоднородные морфоло-
Таблица 10
|
|||||||||
|
Динамика
застойного венозного давления при ритмическом растяжении икроножной
мышцы грузом 3 кг после ее j кураризации, мм рт. ст.
|
|||||||||
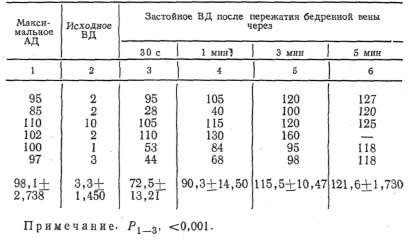 |
|||||||||
|
60
|
|||||||||
|
I
|
||||||
|
гически и
функционально и разряжающиеся с различной частотой, а расслабление
(дилатация) вызывается раздражением проприорецепторов путем их
растяжения вследствие перемещения сустава или сокращения
мышц-антагонистов. Если исходить из обратного — при сокращении мышц не
все проприорецепторы возбуждаются одновременно, то. выходит, что при
растяжении не все мышечные волокна растягиваются и ' возбуждаются
одновременно.
В организме
вся поперечно-полосатая мускулатура находится в состоянии тонуса,
который представляет собой асинхронные сокращения мышечных волокон или
отдельных их групп. При осуществлении какой-либо мышечной работы и
даже в покое скелетные мышцы несколько растянуты, и уже исходя из
этого состояния они сокращаются. Но если мышца растянута, то в ней
растет напряжение, т. е. повышается тонус, что приводит к усилению
асинхронных колебаний мышечных волокон.
Таким
образом, не только при ритмическом растяжении, но и в состоянии покоя
мышц присутствуют необходимые элементы для реализации вибрационного
механизма и, следовательно, проявления микронасосного их свойства.
Кроме этого,
в действительности расслабление мышцы почти всегда сопровождается ее
растяжением. Работами К- Edman, F. Flitney (1977), К- Edman, F.
Flitney (1978), F. Flitney, D. Horst (1978) показано, что при
расслаблении полусухожильной мышцы лягушки некоторые сегменты
мышечного волокна сокращаются, другие удлиняются, а на некоторых
участках имеются группы как укорачивающихся, так и удлиняющихся
саркомеров. Эти же авторы подтвердили зависимость продолжительности
механической активности от длины мышечного волокна.
Таким
образом, свойство, растяжимости принадлежит не только всей мышце, но и
отдельным мышечным волокнам. Следовательно, механизм расслабления
мышцы, сопровождающегося растяжением как на макро-, так и на
микроуровне, весьма сходен с механизмом ее сокращения.
Отсюда
вытекает, что, по всей вероятности, в основе механизма
присасывающе-нагнетательной функции скелетных мышц как при их
сокращении, так и растяжении лежат асинхронные вибрирующие действия
мышечных
|
||||||
|
61
|
||||||
|
волокон, способствующие равномерному их кровоснабжению, а также оказывающие механический эффект на гемодинамику организма.
ЭЛЕКТРОМИОГРАФИЧЕСКИЕ ИССЛЕДОВАНИЯ
Из данных
литературы известно, что пассивное растяжение скелетной мышцы вызывает
в ней появление токов.действия. Их возникновение носит рефлекторный
характер и вызывается раздражением проприорецепторов. При ритмическом
механическом раздражении проприорецепторов в экстрафузальных волокнах
скелетной мышцы появляются рефлекторные биотоки, синхронные с
частотой раздражения. Причем нейромоторные единицы вследствие своей
разнородности возбуждаются не все одновременно, и с увеличением силы
раздражения число одновременно работающих единиц увеличивается.
Путем
ритмического растяжения и раздражения мышечных веретен, воспринимающих
продольное растяжение, Григорьев (1969) показал, что соответственно
частоте ритмического растяжения по рефлекторной дуге возбуждение с
проприорецепторов передается на мышцу и в сокращенных волокнах
возникает ритмическая импульсная активность. После перерезки
седалищного нерва биоэлектрический эффект при пассивном растяжении
исчезает (Уфлянд, 1956). С увеличением напряжения мышцы суммарная
биоэлектрическая активность возрастает (Персон, 1969 и др.).
Работами С.
Hunt, D. Ottoson (1977) установлено, что с увеличением амплитуды
синусоидальных растяжений изолированных мышечньгх веретен кошки
амплитуда рецепторных потенциалов первичных окончаний при постоянной
частоте растяжений возрастает нелинейно.
Для доказательства асинхронизма растяжения мышечных волокон скелетной мышцы использовалась электромиография.
Регистрация
электромиограммы проводилась при нагрузках 3, 6, 8, 10 кг. Выявлено,
что при ритмическом растяжении в мышце возникает импульсация, частота и
амплитуда которой зависят от силы растяжения. При увеличении нагрузки
на мышцу эти показатели увеличиваются. При грузе 3 кг амплитуда
130+2,330 мкв, частота 41—44+2,785 кол/с; при 6 кг — 210+4,480 мкв,
65+3,537 кол/с; при 8 кг —330±1,541 мкв, 136+1,469
|
||||
|
кол/с; при 10 кг амплитуда 440±4,1Ю мкв, частота 87+ ±2,055 кол/с.
При больших
силах растяжения наблюдается возрастание суммарной биоэлектрической
активности (рис. 11), что приводит к усилению
присасывающе-нагнета-тельной функции скелетных мышц.
Таким
образом, электромиографические исследования свидетельствуют о том, что
мышечные волокна растягиваются неодновременно, а при увеличении силы
растяжения количество активных мышечных волокон возрастает.
На основании
полученных данных можно предположить, что механизм
присасывающе-нагнетательной способности скелетных мышц при растяжении
является биофизическим, заключенным в самой скелетной мышце и
совершающемся на уровне микроциркуляции. Он требует
|
|||||||||
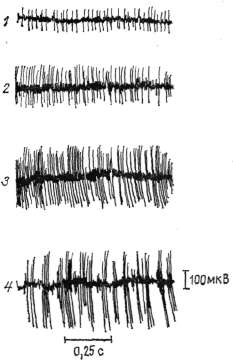 |
|||||||||
|
Рис. 11. Электромиограмма при различной нагрузке ритмического
растяжения: / — ЭМГ при 3 кг ритмического растяжения, 2 — ЭМГ
при 6 кг ритмического растяжения, 3 — ЭМГ при 8 кг ритмического
растяжения, 4 — ЭМГ при 10 кг ритмического растяжения
|
|||||||||
|
63
|
|||||||||
|
дальнейшего,
более детального изучения. По нашим дан ным, этот микронасосный
механизм может найти объяс нение с позиций вибрационной гипотезы,
согласно кото рои мышечные волокна асинхронно, в данном случае не
сокращаясь, а растягиваясь, периодически давят ня кровеносные сосуды
(преимущественно капилляры) чя ключенные в мышце, и тем самым
осуществляют активное проталкивание крови по внутримышечным сосудам
Результаты этих исследований могут использовать™ для совершенствования
существующих и создания но вых.приборов электростимуляции скелетных
мышц и оолее широкого их применения в клинической практике и спортивной
медицине с целью улучшения кровоснаб жения организма и предотвращения
атро|ических яв" лении скелетных мышц. р^ических яв-
|
|||||
|
т
|
|||||
|
Глава VII
|
||||||||
|
ЭФФЕКТИВНОСТЬ МИКРОНАСОСНОЙ
ФУНКЦИИ СКЕЛЕТНЫХ МЫШЦ
ПРИ РАСТЯЖЕНИИ ПОСЛЕ ОДНО-
И ТРЕХМЕСЯЧНОЙ ФИЗИЧЕСКОЙ
ТРЕНИРОВКИ
Существование
микронасосного свойства скелетных мышц при их растяжении поставило
вопросы о том, как оно реагирует и в какую сторону изменяется под
влиянием различного объема двигательной активности и при гипокинезии.
Изучение
микронасосной деятельности скелетных мышц собак, подвергавшихся одно- и
трехмесячной тренировке в беге на третбане, проводилось путем
преграждения венозного оттока при сохраненном артериальном притоке
крови к мышце, находящейся в условиях физиологического покоя, и при
ритмическом растяжении ее различной силой.
Застойное
венозное давление икроножной мышцы в условиях покоя у собак, в течение
одного месяца тренировавшихся на третбане, повышалось, но к концу 10-й
минуты все же не достигало уровня максимального артериального давления
(табл. 11). Оно было несколько выше (94±4,9), чем у нетренированных
животных (84,3±4,9 мм рт. ст.), однако достоверных различий между ними
не выявлено.
После
трехмесячной тренировки застойное венозное давление в условиях
физиологического покоя икроножной мышцы повышалось более стремительно,
достигая 101,4±5,035 мм рт. ст., но и в этом случае оно не превышало
уровень максимального артериального давления (табл. 12).
Если
сравнить данные, полученные на собаках до, а также после одно- и
трехмесячной тренировки, то можно -увидеть, что застойное венозное
давление имеет тенденцию к увеличению: у нетренированных собак оно
составило 84,3±4,96 мм рт. ст., после одномесячной тренировки — 94,0
±4,932, а после трехмесячной—101,4±5,035 мм рт. ст., причем застойное
венозное давление после трехмесячной тренировки достоверно превышало
таковое у '. нетренированных.
|
||||||||
|
5. Зак. 837
|
||||||||
|
65
|
||||||||
|
Таблица 11
Влияние одномесячной
тренировки на динамику застойного венозного давления при растяжении
Икроножной мышцы собак, находящейся в условиях физиологического покоя,
мм рт. ст.
|
||||||||
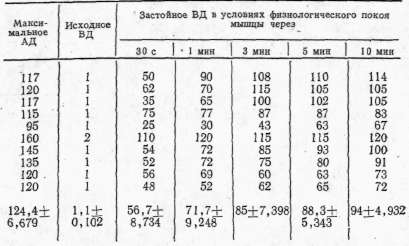 |
||||||||
|
Измерение тонуса икроножной мышцы у собак стоя показало, что в процессе тренировки он возрастает с 91,9±4,110 г/см2 до 106,75±3,974 г/см2, а в положении лежа, т. е. при расслаблении, наоборот, снижается с 53,5±4,110 г/см2 до 38,25±3,725 г/см2
(табл. 13). Тонус же изолированной икроножной мышцы у тренированных
животных с высокой достоверностью (Р<0,001) больше, чем у
нетренированных.
Следовательно,
физическая тренировка приводит к усилению тонуса мышц, т. е. к
повышению адаптационных возможностей мышечной ткани, что хорошо
согласуется с данными литературы (Ефимов, 1947; Кесарева, 1960;
Фарфель, 1960 и др.). По мнению многих авторов (Rohracer, 1949, 1955,
1959; Махмудов, 1965; Жуков, 1969 и др.), тонус скелетных мышц
поддерживается за счет асинхронных сокращений мышечных волокон или их
групп, возникающих попеременно в различных местах мышцы. В основе
микронасосной деятельности скелетных мышц, по нашим представлениям,
также лежат асинхронные сокращения мышечных волокон, которые
периодически давят -на цитомембрану эндотелиоцитов капилляра, в
результате чего его просвет то уменьшается, то увеличивается. Эти
механические взаимодействия мы-
66
|
||||||||
|
Таблица. 12
|
||||||||||||
|
Влияние трехмесячной физической тренировки на динамику
застойного венозного давления при растяжении икроножной
мышцы собак,' находящейся в условиях физиологического покоя,
мм рт. ст.
|
||||||||||||
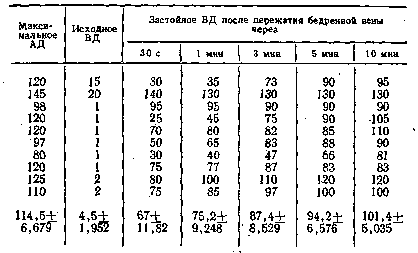 |
||||||||||||
|
Та блица 13
Изменение тонуса икроножной мышцы собак под влиянием трехмесячной тренировки, г/см2
|
||||||||||||
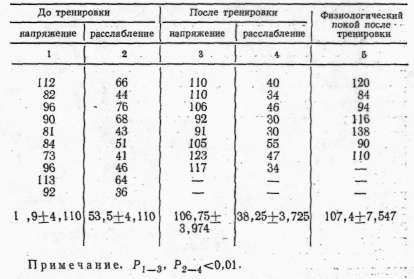 |
||||||||||||
|
67
|
||||||||||||
|
шечных
волокон с капиллярами имеют волнообразный характер и образуют своего
рода эндотелиальномышеч-ный комплекс, который выполняет функцию
микронасосов, проталкивающих .кровь на каждом данном участке
мик-роциркуляторного русла (Аринчин, Володько, 1979). Достоверное
увеличение застойного венозного давления у тренированных животных по
сравнению с нетренированными связано с повышением тонуса икроножной
мышцы
ммрпгст. 300 л
|
||||||||||||
 |
||||||||||||
|
Рис. 12.
Влияние физической тренировки на зависимость повышения застойного
венозного давления от силы растяжения: / — после трехмесячной
тренировки, 2 — после одномесячной тренировки, 3 — у нетренированных собак
|
||||||||||||
|
в процессе
тренировки. Усиление микронасосной деятельности скелетных мышц, даже в
условиях физиологического покоя, после тренировки, по всей
вероятности, можно объяснить увеличением количества асинхронно
сокращающихся мышечных волокон, а также их взаимодействием с
внутримышечными сосудами, в основном капиллярами.
Физическая
тренировка собак в течение одного месяг ца вызывала усиление
присасывающе-нагнетательной функции икроножной мышцы. Застойное
венозное давление при нагрузках 1, 2, 6, 10 кг достоверно увеличивало
уровень такового у нетренированных собак. Причем уже при малых
растягивающих силах наблюдалось превышение застойным венозным
давлением уровня максимального артериального (рис. 12).
После
физической тренировки в течение трех месяцев застойное венозное
давление при различных силах растяжения икроножной мышцы достоверно
(Р<0,001) превышало таковое как у нетренированных собак, так и после
одномесячной тренировки.
По показателям застойного венозного давления в условиях растяжения можно косвенно судить и об изме-
|
||||||||||||
|
68
|
||||||||||||
|
нении
динамического тонуса, который определяется сопротивлением мышцы в
продольном направлении (Кесарева, 1960). По нашим данным, он
повышается. Это является подтверждением данных о том, что тренировка
способствует усилению микронасосной функции при растяжении, вероятно,
в связи еще и с улучшением растяжимости самой мышцы.
Следовательно,
физическая тренировка есть тренировка и микронасосного механизма не
только при сокращении, но и растяжении скелетных мышц. Она приводит к
тому, что скелетные мышцы более эффективно помогают сердцу в
продвижении крови по большому кругу кровообращения. Однако мы изучали
влияние только одно-и трехмесячной тренировок на
•присасывающе-нагнета-тельную способность скелетных мышц. Возможно, что
очень длительные тренировки (6,12 мес и больше), осуществляемые с
большой интенсивностью, могут привести не к увеличению, а, наоборот,
вследствие перетренированности к снижению эффективности внутримышечных
периферических «сердец». Продолжение исследований в этом направлении
чрезвычайно важно для тренеров, которые готовят спортсменов
международного класса, тренирующихся на грани функциональных
возможностей организма.
|
||||
|
держанию и
восстановлению сердечной деятельности (Шушков, 1950). Восстановление
сердца с помощью данного метода совершается, по-видимому, вследствие
возможного растяжения миокарда и ритмического по механизму
гемодинамического удара проталкивания крови по коронарным сосудам,
приводящим к нормализации кровоснабжения миокарда.
Все
изложенное проливает свет на весьма противоречивые мнения ученых о
том, как влияют сокращения мышечных волокон и слоев миокарда на
заключенные в них кровеносные сосуды и кровоснабжение всего
работающего сердца в норме и патологии. Вопреки мнению о том, что
экстр аваскулярное сжатие внутримиокардиальных сосудов препятствует
кровоснабжению сердца, получены данные, подтверждающие, что как при
сокращении (систоле), так и расслаблении (диастоле) совершается
проталкивание крови по микрососудам миокарда и выталкивание крови в
сторону синуса, что, несомненно, способствует кровоснабжению ткани
сердца. В этом состоит не отрицательная, а положительная роль
экстраваску-лярного сжатия в кардиогемодинамике.
Если бы
этого механизма не существовало, то сердечная ткань лишалась бы
некоторой доли кровоснабжения. Следует заметить, что выталкиваемая
кровь при систоле по объему меньше, чем та, которая протекает по
коронарным, сосудам в фазу диастолы и паузы, но она выбрасывается с
силой, превышающей исходное давление.
Сердце,
таким образом, работает на два фронта: внешний — для кровоснабжения
всего организма и внутренний — для кровоснабжения самого себя. Этот
внутренний механизм можно образно назвать как «сердце в-сердце»,
«работающий сам на себя». Это и понятно, ибо трудно допустить, чтобы
столь мощная энергия миокарда тратилась только на внешнюю работу, а по
отношению к самому себе была бы помехой. Природа, создавшая все
живое, в том числе и человека, не могла использовать сократительный
механизм миокарда только на внешнюю работу в ущерб внутренней, а стала
сочетать,, как легко в этом убедиться, и то и другое с положительным
эффектом. Миокард, как и мышца, «холостого хода» не имеет, ибо
проявляет насосную способность как при сокращении, так и расслаблении,
растяжении.
Все эти данные целесообразно шире использовать в профилактической и клинической практике.
|
||||
|
Глава IX
|
||||||||
|
О МЕХАНИЗМАХ АКТИВНОСТИ
МИКРОНАСОСОВ
ПОПЕРЕЧНО ПОЛОСАТЫХ ТКАНЕЙ
В УСЛОВИЯХ ФИЗИОЛОГИЧЕСКОГО ПОКОЯ
|
||||||||
|
Присасывающе-нагнетальное
свойство присуще микронасосам скелетных мышц и миокарда не только при
сокращении, но и растяжении. Это всеобщее свойство всех
поперечно-полосатых тканей. Но как скелетные мышцы, так и миокард с
точки зрения трехфазной структуры сердечного цикла пребывают не только
в состоянии сокращения и расслабления, растяжения, но и в состоянии
той или иной степени физиологического покоя. И если согласиться с тем,
что состояние физиологического покоя в функционировании
поперечно-полосатых мышечных тканей (скелетной и сердечной)
существует, то возникает вопрос о том, как они в этом состоянии
снабжаются кровью. Нельзя отрицать, что это осуществляется с помощью
механизма сужения и расширения, т. е. изменения просвета и проходимости
сосудов, механизмов вазомоторики, регулируемой нервными и гуморальными
механизмами.
Но возникает
другой вопрос, касающийся обнаруженного микронасосного механизма,
который заключен в этих тканях. Вопрос о том, как ведет себя этот
микронасосный механизм, когда скелетные мышцы и миокард находятся в
покое, бездействует он или же как-то себя проявляет, а если да, то
как. Каким образом методически можно приблизиться к изучению этого
вопроса, чтобы ответить на него?
Задача
чрезвычайно сложная, трудная и на первых порах представляется
выполнимой с использованием препаратов, в частности, кураре,
блокирующих передачу импульсов с нервов на мышцу. Под влиянием этого
препарата снижаются вибрационные колебания мышечных волокон. А при
пережатии бедренной вены, идущей от икроножной мышцы, застойное
венозное давление при всех прочих равных условиях поднимается
значительно медленнее и на меньшую высоту по сравнению с конт-
|
||||||||
|
87
|
||||||||
|
рольной мышцей, у которой сохраняется нормальная передача импульсов с нервов на мышцу.
Блокирование
импульсов и снижение скорости и высоты подъема застойного венозного
давления — объективный показатель того, что и в покое микронасосный
механизм все же работает, но он проявляет себя в меньшей степени по
сравнению с сокращением и растяжением скелетной мышцы и миокарда.
О
существовании микронасосной функции скелетных мышц в физиологическом
покое убеждают опыты о зависимости таковой от тонического напряжения
мышечной ткани. В них установлено, что у собак с более выраженным
тонусом икроножной мышцы, находящейся в состоянии физиологического покоя, порядка 74—92: г/си2 застойное
венозное давление повышалось более стремительно и достигало больших
величин, чем у собак со слабо выраженным тонусом в покое, равным 44—60
г/см2.
В дополнение
к этим экспериментам проведены серии исследований с применением метода
электромиографии (ЭМГ), которые также подтверждают наличие
микронасосной функции скелетных мышц в покое. Максимальное
артериальное давление в покое составляло 115± ±7,8 мм рт. ст., а
застойное венозное — 78±'3,7 мм рт. ст. После введения диплацина через
20 мин застойное венозное давление уменьшилось до 62±3,3 мм рт. ст.
При растяжении же скелетной мышцы грузом 3 кг оно увеличилось до 121
±1,7 мм рт. ст. и стало превышать максимальное артериальное (115±7,8
мм рт. ст.). Следовательно, микронасосы, заключенные в скелетной
мышце, работают и в покое, но диплацином угнетаются, растяжение же их
стимулирует, несмотря на предшествующее .блокирование микронасосного
механизма этим препаратом.
Все это
убедительные доказательства реальности работы микронасосов скелетных
мышц даже в покое, ибо они не только перфузируются, но и
самообеспечиваются кровью. Вибрационный механизм скелетных мышц
действует и в покое. А если это объективная реальность, то ее можно
наблюдать во всех других адекватных условиях. •■■■■<
В связи с
обнаружением микронасосного свойства скелетных мышц и миокарда,
проявляющегося даже в покое, открываются новые, ранее неизвестные
возможности и проблемы изучения структуры и функции ске-
88
|
||||
|
•летных мышц
как самостоятельных- гемодинамических насосов,
присасывающе'-нагнетательная деятельность которых совершается на
микроуровне в масштабах двигательных единиц скелетных мышц и миокарда.
Двигательная
единица (ДЕ) — это пучок, состоящий из 100 и более мышечных волокон,
покрытый соединительной тканью и пронизанный капиллярами, которые от
поперечно расположенного по отношению к мышце артериального сосуда
идут параллельно мышечным волокнам в обе стороны и образуют
своеобразные эндотели-•ально-мышечные комплексы — самостоятельные
субъединицы (микронасосы), представленные в каждой ДЕ в огромном
количестве.
Знать, как
устроены эти насосы и как действуют, очень важно для науки и практики,
для построения правильного образа жизни, поддержания деятельности
периферических «сердец» на должном уровне с целью оказания
повседневной помощи сердцу и гарантирования его долгой бесперебойной
работы. Чрезвычайно важно также правильное развитие всех этих
механизмов у детей, начиная от рождения.
: Строение и
деятельность микронасосов скелетных мышц и миокарда полезно учитывать
при спортивной тренировке с целью достижения наивысших мировых
спортивных результатов. Они могут использоваться при прогнозировании и
отборе спортсменов для того или иного вида спорта, а также для
предупреждения перегрузок, перетренировок и их реабилитации.
Эти знания
необходимы для профилактической и клинической медицины, для дальнейшей
разработки теорий и практики нарушений нейромоторного и микронасосного
аппаратов и механизмов скелетных мышц как периг ферических «сердец»,
активных помощников сердца человека.
С помощью
прикрепляемых к коже (например, руки человека) датчиков
гемомиодинамографа, сконструированного и созданного в лаборатории
кровообращения Института физиологии АН БССР и СКВ Белорусского
политехнического института, графически регистрируются электрические
биопотенциалы мышц в виде электромио-граммы (ЭМГ), вибрационные
колебания мышечных волокон в виде вибромиограммы (ВМГ) и звуковые
колебания этих же мышечных волокон в виде фономиограм-мы (ФМГ). •;..--. '.:..::■
|
|||||
|
89
|
|||||
|
Оказалось,
что при волевом, произвольном напряжении скелетных мышц предплечья
человека микронасосная их эффективность находится в прямой зависимости
от амплитуды и частоты электрических, звуковых и вибрационных
колебаний мышечных волокон и их двигательных единиц —- мионов
(Ермолова, 1971).
Правда,
оставался труднообъяснимый факт резкого усиления кровотока из скелетной
мышцы после прекращения длительного, тетанического ее напряжения. При
этом ЭМГ не регистрировалась и надо было полагать, что биоэлектрическая
активность отсутствовала. Следовательно, по этому показателю
микронасосы в скелетных мышцах не работали, однако кровь вытекала из
мышцы в максимальном количестве.
На первый
взгляд это говорит не в пользу вибрационной гипотезы деятельности
внутримышечных периферических «сердец» (ВПС), и усиление их
кровоснабжения, в данном случае, совершается якобы по механизму
вазодилатации. Но это только на первый взгляд, ибо звуковые (ФМГ) и
вибрационные (ВМГ) колебания мышечных волокон продолжали
регистрироваться. Они медленно затухали, а вместе с ними снижалась и
так называемая постконтракционная гиперемия (Ермолова, 1971), которую
некоторые исследователи объясняли только постконтракциоцным расширением
сосудов. Но если принимать во внимание зависимость ее от звуковых и
вибрационных колебаний мышечных волокон, то следовало бы считать, что
и она есть не постконтракционная гиперемия, а наивысшая микронасосная
эффективность, проявляющаяся вибронасосным механизмом мышц в условиях
снятия напряжения, сокращения мышцы и, сле-довально, внешнего давления,
затрудняющего нагнетательную функцию в этих условиях самих
микронасосов.
При этом,
однако, возникают такие очень важные вопросы: почему и как могли
звучать и продолжать вибрировать мышечные волокна или мионы ДЕ, если
импульсы, идущие к мышцам из центральной нервной системы (из
головного мозга) сняты? Могут ли мышцы вибрировать без поступающих к
ним импульсов? Способны ли они к спонтанной вибрации?
Лишь четкие,
убедительные ответы на эти вопросы могут служить объяснением данного
феномена. Что можно привести в пользу сказанного? Житейские наблюдения
о том, что у длительно и много физически и напря-
|
|||||
|
90
|
|||||
|
женно
работающих лиц наступает утомление, они стремятся отдохнуть и когда
лягут, то говорят, что их мышцы «гудят». Из головного мозга им не
поступают приказы работать, однако субъективные ощущения таковы, что
скелетные мышцы после таких напряжений продолжают вибрировать,
«гудеть». Но это всего лишь субъективные ощущения. А о чем говорят
объективные научные данные?
С помощью
селективных электродов, позволяющих отводить биопотенциалы от немногих,
например трех ДЕ (Саплинскас, 1970—1980), обнаружено, что в покое
регистрируется спонтанная электрическая активность ДЕ низкого
вольтажа, которая выявляется на осциллоскопе при больших усилениях и
проявляется в незначительных колебаниях активности. Частота их
колеблется от 13 до 25 Гц. Механизм, способствующий спонтанному
регулярному разрешению двигательной концевой пластинки (ДКП),
неизвестен и требует специального изучения '(Buchthal, Rosenfalck,
1966).
Любопытно^,
что спонтанная электрическая активность регистрируется в двигательных
концевых пластинках, в мышечных волокнах (MB) как перед, так и после
утомительных нагрузок. Спонтанная электрическая активность мышечных
волокон у человека регистрируется и при их денервации, в частности при
повреждении нервов у спортсменов. Эти спонтанные потенциалы низкой
амплитуды с начальным отрицательным отклонением длительностью от 0,5
до 3 мс М. Stohr (1977) относит к «миниатюрным потенциалам» ДКП,
которые впервые были описаны Ф. Фатом и Б. Катцом (Fatt, Katz, 1952).
Следует
подчеркнуть, что электрическую активность ДКП можно отвести двумя
игольчатыми электродами с расстоянием между ними не более 0,4 мм, так
как с увеличением этого расстояния биопотенциалы не регистрируются.
Следовательно, волна деполяризации и биоэлектрической активности
распространяется в очень узких пределах и другими методами она не
обнаруживается. По-видимому, вот почему на электромиограмме ничего не
видно, а ДЕ сокращаются, и, следовательно, микронасосы даже в этом
состоянии все же работают.
Усиление
спонтанной активности ДКП, разряжающейся асинхронно с большой
вариабельностью межимпульсных интервалов, по мнению Ю. С. Саплинскаса
(1984), следует рассматривать как физиологическое
|
|||||
|
S1
|
|||||
|
явление. Оно в сопоставлении с нашими исследованиями способствует работе микронасосов скелетных мышц даже в покое. Однаког
по мнению того же автора, одинаковой длительности последовательные
межимпульсные интервалы спонтанно разряжающейся ДКП в ряде случаев
следует рассматривать как один из критериев перегрузки нервно-мышечной
системы, ведущей к патологий. И:с нашей точки зрения, нарушение
асинхронности колебаний мышечных волокон свидетельствует о перегрузке,
переутомлении, что тоже может привести к патологии.
У спринтеров
превалируют мышечные волокна II типа. Вероятно, они предопределены
генетически и успешно развиваются при тренировке, под влиянием которой
становятся толще, сила их возрастает и сокращаются они быстрее. Эти
быстрые волокна богаче гликогеном, но беднее капиллярами, митохондриями
по сравнению с медленными волокнами I типа, а условия неутомляемости и
кровотока их совпадают (Замостьян, 1976).
Ю. С.
Саплинскас (1980) полагает, что спонтанная электрическая активность
мышечных волокон не всегда означает денервацию. В. Салтин (Saltin et
al., 1977) считает, что причиной появления спонтанных осцилляции покоя
может быть потеря контактов мышечных волокон с двигательным нервом при
утомительных физических нагрузках, а затем этот контакт
восстанавливается.
Регистрируемые
на ЭМГ спонтанные потенциалы с амплитудой 100—150 мкВ и частотой 6—-12
Гц, согласно Ю. С. Саплинскасу (1984), следует отнести не к
фибрилляциям, а к фасцикуляциям.
Утомительная
физическая работа вызывает спонтан-ную электрическую активность ДКП и
появление нерегулярных, а также регулярных спонтанных осцилляции типа
фасцикуляций, а иногда и фибрилляций, что служит предупреждением
нарушений функционирования мышечных волокон и двигательных центров и
может носить временный, преходящий характер.
При
денервации появляются спонтанные потенциалы в некоторой степени сходные
с теми, которые возникают при утомлении от физических нагрузок с
сохраненной иннервацией мышечных волокон.
Приведенные
фактические данные убедительно свидетельствуют о том, что
микронасосный механизм скелетных мышц, а вероятно, и миокарда
функционирует и в состоянии их физиологического покоя.
|
||||
|
ЗАКЛЮЧЕНИЕ
|
||||||
|
Обнаружение
микронасосного свойства поперечнополосатой ткани, формирующей
скелетные и сердечную мышцы, имеет принципиальное значение как с
научной, так и с практической точки зрения. На основании этих новых
фундаментальных научных данных скелетные мышцы и миокард должны
рассматриваться не только как потребители крови, нагнетаемой сердцем, и
перфу-зируемые органы, величина перфузии которых зависит от изменений
тонуса и просвета сосудов, но и как физиологические вибраторы и
самообеспечивающиеся органы. Этим они обязаны заключенному в них
микронасосному механизму, выполняющему две функции: внутреннюю
—-самообеспечивание кровью скелетных мышц и миокарда и внешнюю —
присасывание артериальной крови, проталкивание ее по внутримышечным
капиллярным сосудам и нагнетание венозной крови с потенциальной силой,
превышающей максимальное артериальное давление. Скелетные мышцы — это
своеобразные периферические «сердца», которые вместе с другими
экстракарди-альными насосами — грудным, брюшным, диафрагмаль-ным, а
также венозными помпами помогают сердцу человека и животных. Без них
оно не в состоянии обеспечить нормальную циркуляцию крови по большому
кругу кровообращения и кровоснабжение организма.
На основе
этих данных должен быть уточнен взгляд на рабочую гиперемию скелетных
мышц, усиление кровоснабжения которых при работе объяснялось падением
тонуса и расширением сосудов. Следует подчеркнуть, что само
явление перфузии, конечно, не исключается, но в работе скелетных мышц
оно занимает весьма незначительное место. В связи с этим становится
очевидным тот факт, что ни одной из многих гипотез рабочей
гиперемии расширения внутримышечных сосудов, в том числе и
ги-стомеханической гипотезой вазодилатации В. М. Хаюти-на, невозможно
объяснить присасывающе-нагнетатель-ную способность скелетных мышц.
|
||||||
|
93.
|
||||||
|
Если
скелетной мышце присуща только перфузия, то по закону сообщающихся
сосудов невозможно превышение давлением на выходе из мышцы давления на
ее входе. Нет ответа на многие другие вопросы, в частности почему при
денервации, а также иммобилизации мышц (например, заключенных под
гипсом, даже с сохраненной иннервацией) они атрофируются. Сердце
по-прежнему посылает кровь всем органам в том же количестве, но если
работающие мышцы продолжают функционировать нормально, то
иммобилизованные почему-то атрофируются. Как эти, так и многие другие
факты непонятны с точки зрения рабочей гиперемии за счет вазодилатации,
перфузии, а с позиции вибрационной гипотезы и микронасосного механизма
они становятся вероятными и объяснимыми. Если микронасосы мышц
работают, они могут нагнетать венозную кровь с превышением
максимального артериального давления. Но когда микронасосы перестают
работать, не снабжают мышечную ткань питательными веществами и
кислородом, то мышцы атрофируются.
Как показано
в данной работе, микронасосы в какой-то степени продолжают работать и в
состоянии покоя и даже после блокирования передачи нервных импульсов,
побуждающих мышцу к работе, но этот уровень их деятельности
недостаточен для того, чтобы поддерживать длительно скелетную мышцу в
нормальном функциональном состоянии, а тем более обеспечивать ее
дальнейшее развитие и совершенствование, и она атрофируется. В
условиях сниженной двигательной активности — гипокинезии микронасосы
угнетаются вплоть до прекращения их действия, в чем состоит одно из
губительных ее последствий, и наоборот, развиваются и совершенствуются
они при оптимальной по объему и интенсивности физической активности,
особенно тренировке, усиливаются при массаже, проявляются при
холодовой мышечной дрожи и т. д. Но при всем этом оставался открытым
вопрос, действует ли микронасосный механизм при растяжении скелетных
мышц, а также при расслаблении миокарда, диастоле и паузе. Если он
сохраняется и продолжает работать, то в каких параметрах он
проявляется и что влияет на него? Эти знания необходимы для выяснения
процессов, лежащих в основе растяжения мышц, и управления ими.
В данной работе приведены экспериментальные до-
|
|||||
|
94
|
|||||
|
казательства
того, что скелетная мышца ив условиях ритмического растяжения проявляет
четко выраженную микронасосную функцию. Наряду с нагнетательной
насосной деятельностью скелетная мышца в условиях растяжения обладает
и присасывающей способностью, которая выражается в понижении
артериального давления вплоть до отрицательных величин на фоне
пережатия артерии и вены. Эффективность присасывающе-нагнета-тельной
функции находится в прямой зависимости от силы растяжения в диапазоне
от 1 до 10 кг. Она зависит также от частоты и амплитуды
электромиографической активности скелетной мышцы, характеризующей
состояние ее микронасосного механизма.
Результаты
исследований, полученные при денерва-ции, а также после кураризации
скелетной мышцы, свидетельствуют о том, что
присасывающе-нагнетательная функция обусловлена главным образом
биомеханическими воздействиями мышечных волокон на внутримышечные
сосуды, в основном капилляры.
Ритмически
растягиваемая полностью изолированная скелетная мышца способна
обеспечивать движение крови по искусственному замкнутому кругу
кровообращения и проявлять себя как самостоятельное периферическое
«сердце», эффективное при растяжении мышц.
Полученные
данные раскрывают новые стороны деятельности скелетных мышц не только
при сокращениях, но и их растяжении и созвучны с некоторыми сторонами
замечательного открытия А. Н. Студитского — регенерации скелетных мышц
и созданного им на этой основе учения. «Сокращаясь,— писал он,— мышца
разрушается подобно любой работающей машине. Расслабляясь, мышца, в
отличие от машины, восстанавливает разрушенное. Этот процесс
контролируется трофическим действием нерва» (Студитский, 1983, с. 32).
Содержащиеся
в настоящей книге новые сведения о проявлении микронасосного свойства
скелетных мышц объясняют механизм протекания восстановительных
процессов в скелетных мышцах с гемодинамической точки зрения. Они
совершаются при расслаблении, потягивании и растяжении скелетных мышц
под влиянием не только трофических нервов, но и активации
присасываю-ще-нагнетательной деятельности микронасосов с усилением
самообеспечения, питания мышц кровью.
А. Н. Студитский (1980) рассматривает также меха-
|
|||||
|
95
|
|||||
|
низмы тесной
взаимосвязи между деятельностью мышц и мозга, осуществляющиеся с
помощью гипотетических вешеств миотрофинов и нейротрофинов. -
Взаимосвязь между мышцами и мозгом поддерживается и совершенствуется
благодаря помощникам сердца и главным образом скелетным мышцам как
перифериче--ским «сердцам», работающим и при растяжении. Они подают
венозную кровь и содержащиеся в ней миотро-фины к сердцу для его
лучшего наполнения в фазах диастолы и паузы. Сердце же в свою очередь
больше нагне--тает крови всему организму и, конечно, головному мозгу,
принося ему с кровью скелетные миотрофины, кислород и питательные
вещества, удаляя ядовитые продукты жизнедеятельности.
И наконец,
необходимо подчеркнуть, что микронасосная способность принадлежит
также миокарду не только при его сокращении и массаже, но и
расслаблении, растяжении, диастоле и паузе. Эти данные тоже интересны
и важны в свете чрезвычайно важного открытия Л. В. Полежаева (1977) о
регенерации миокарда. Процессы восстановления в миокарде (как в норме,
так и при инфаркте) совершаются не только вследствие доставки крови,
кислорода и питательных веществ по коронарным сосудам, обладающим
вазомоторикой, но и благодаря работе микронасосов миокарда, работающих
как в фазе систолы, так и в фазах диастолы и паузы.
Этот
обнаруженный внутримиокардиальныи микронасосный механизм, образно
говоря, «сердце в сердце», представляет самостоятельный интерес и
требует дальнейшего специального изучения, так как его нарушения могут
оказаться источниками заболеваний сердца. Многие применяемые в
клинической практике сердечные препараты, несомненно, влияют не только
на коронарные сосуды, но и на внутримиокардиальныи микронасосный
механизм, и эти вопросы становятся неотложными для кардиологической
науки и практики.
|
||||
|
ЛИТЕРАТУРА
|
||||||||
|
Аберберг-Ауешканпе Л. А. Показатели
периферического кровообращения при физических нагрузках у лиц разного
возраста: Автореф. дис. ... канд. мед. наук. Рига, 1977. 18 с.
Аденский А. Д. Венозное давление и значение в клинике сердечно-сосудистых заболеваний. Минск, 1953. 215 с.
Алашеева В. М. Особенности
мйкроциркуляторного кровеносного русла и ДНК миокарда животных при
гиподинамии и интенсивной физической нагрузке//Материалы \ Всесоюз. науч. конф. по спорт, морфологии. М., 1975. С. 9—13.
Алферова И. В., Егоров А. Д., Полякова А. П. Динамика фазовой структуры диастолы в 140-суточном космическом полете//Косм, биология и авиац. медицина. 1980, № 5. С. 32—35.
Андреев Ф. А. Диастола
сердца: Критический разбор учения об активно-аспирационной функции
диастолы: Диастолическая аккомодация сердца и роль vis a tergo крови.
М., 1916. 156 с.
Аринчин Н. И. О
функциональном взаимоотношении тонического состояния сосудов и
напряжения скелетной мускулатуры в конечностях человека//Сб. работ II
науч. сес. Гродн. гос. мед. ин-та. Гродно, 1960. С. 62.
Аринчин Н. И. Комплексное изучение сердечно-сосудистой системы. Минск, 1961. 220 с.
Аринчин Н. И. Методика наблюдения за активностью фазы диастолы сердца//Здравоохранение Белоруссии. 1962, № 8. С. 57—59.
Аринчин Н. И.. Кузнецов В. И., Рожко В. А. и др. К вопросу о физиологии и патологии диастолы сердца//Материалы IV Всесоюз. конф. патофизиологов. Тбилиси, 1964. Т. 1. С. 15—17.
Аринчин Н. И. Типы саморегуляции кровообращения // Материалы VI науч. сес. Гродн. -тос. мед. ин-та и Всесоюз. симпоз. по тиамину. Минск, 1966. С. 38—39.
Аринчин Н. И. Эволюционное и клиническое толкование электрокардиограммы и фаз сердечного цикла. Минск, 1966. 224 с.
Аринчин Н. И., Кулаго Г. В. Гипертоническая болезнь как нарушение саморегуляции кровообращения. Минск, 1969. 99 с.
Аринчин Н. И., Сенько Ф. Н. Фазы и периоды сердечного цикла. Минск, 1970. 144 с.
Аринчин Н. И., Калинина Т. В., Логвинов Э. М. Общая закономерность изменений кровообращения в процессе старения человека // Докл. АН БССР. 1971. Т. 15, № 11. С. 1041—1052.
Аринчин И. И., Ермолова А. С. Роль
физических упражнений в развитии механизмов сокращения и кровоснабжения
скелетных мышц//Физическая культура в режиме труда и отдыха: Материалы.
Междунар. конф. М., 1971. С. 12—13.
|
||||||||
|
7. Зак. 837
|
||||||||
|
97
|
||||||||